人工智能架构演化中的普适统计特征 Universal statistical signatures of evolution in artificial intelligence architectures
用 935 次消融实验证明 AI 架构演化与生物演化共享同样的统计规律
前置知识
适应度效应分布 (DFE)
Distribution of Fitness Effects,描述生物群体中新突变对个体适应度影响的概率分布,通常以"有害/中性/有益"三类的比例来刻画,并用 gamma 或对数正态等函数拟合其连续形态。它是连接基因型空间拓扑与可观察演化动态的核心桥梁。
本文的核心被解释变量就是 AI 架构修改的"适应度效应分布",必须先理解 DFE 在生物演化文献中是如何被估计和比较的,才能看懂作者为何把 935 个消融实验折算成相对适应度差 $\Delta$ 并与八种生物做 KS 检验。
消融实验 (Ablation Study)
机器学习中评估单个组件贡献的标准做法:移除或替换模型的一个部件(如某层、某正则项),保持其它超参与训练流程不变,记录下游指标变化。本文中 $\Delta = (\text{ablated} - \text{baseline})/|\text{baseline}|$ 被类比为生物学的选择系数 $s$。
数据集的主体 935 次消融直接来源于 arXiv 论文,这是把"AI 架构"映射为"演化谱系"的唯一可量化操作;不熟悉消融范式就无法理解作者为何要把 140 次手工整理与 795 次 LLM 抽取的样本放在一起。
适应度景观 (Fitness Landscape)
Wright 提出的概念:把基因型映射到一个高维空间中的"高度"函数,演化被视为在景观上的爬山过程。景观的局部崎岖程度决定 DFE 的尾部厚度,全局地形则决定多样化与收敛的宏观模式。
全文的论证链条是"AI 与生物的统计签名一致 → 二者所处适应度景观的拓扑相似 → 演化结构由景观决定而非搜索机制",必须先接受景观这一抽象,才能读懂作者为何把定向工程类比为'有眼力的诱变剂'。
重尾分布与 Student t 分布
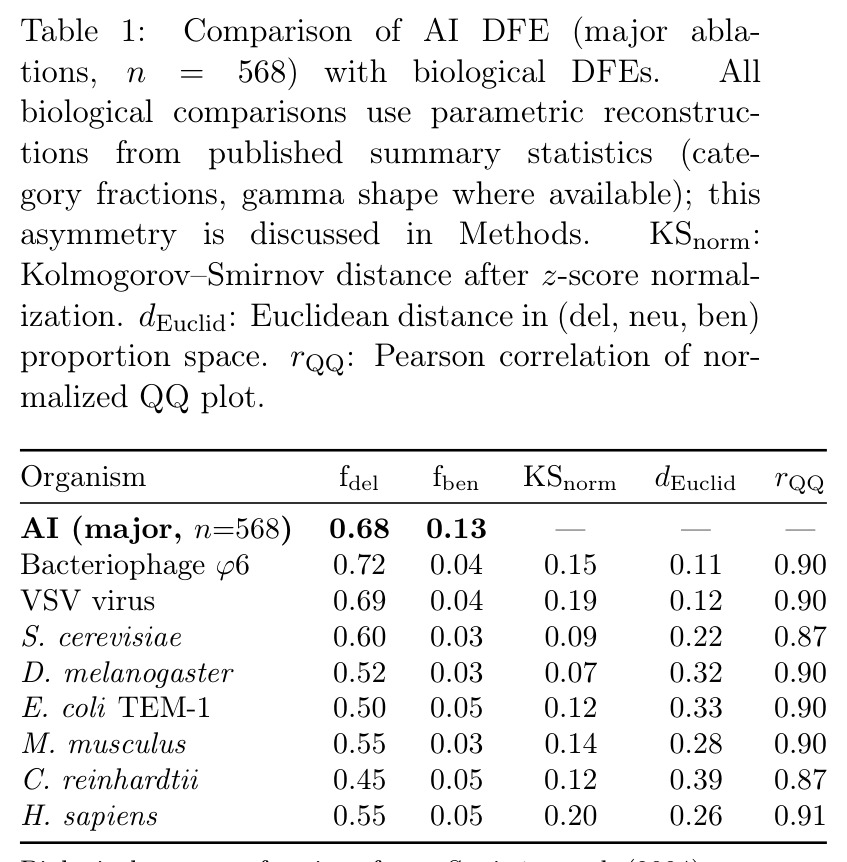
相比正态分布,重尾分布在极端值附近概率更高,Student t 分布通过较小的自由度 $\nu$ 来刻画这种"偶发但显著"的偏离样本均值的现象。本文中 DFE 偏度 $-2.23$、峰度 $29.2$,正是用 Student t(而非正态或 Laplace)来拟合才得到最低 AIC。
理解尾部厚度的生物意义——少数极大正向/负向突变——是判断"AI 演化是生物演化的加速版"这一论断是否成立的关键统计依据。
收敛演化 (Convergent Evolution)
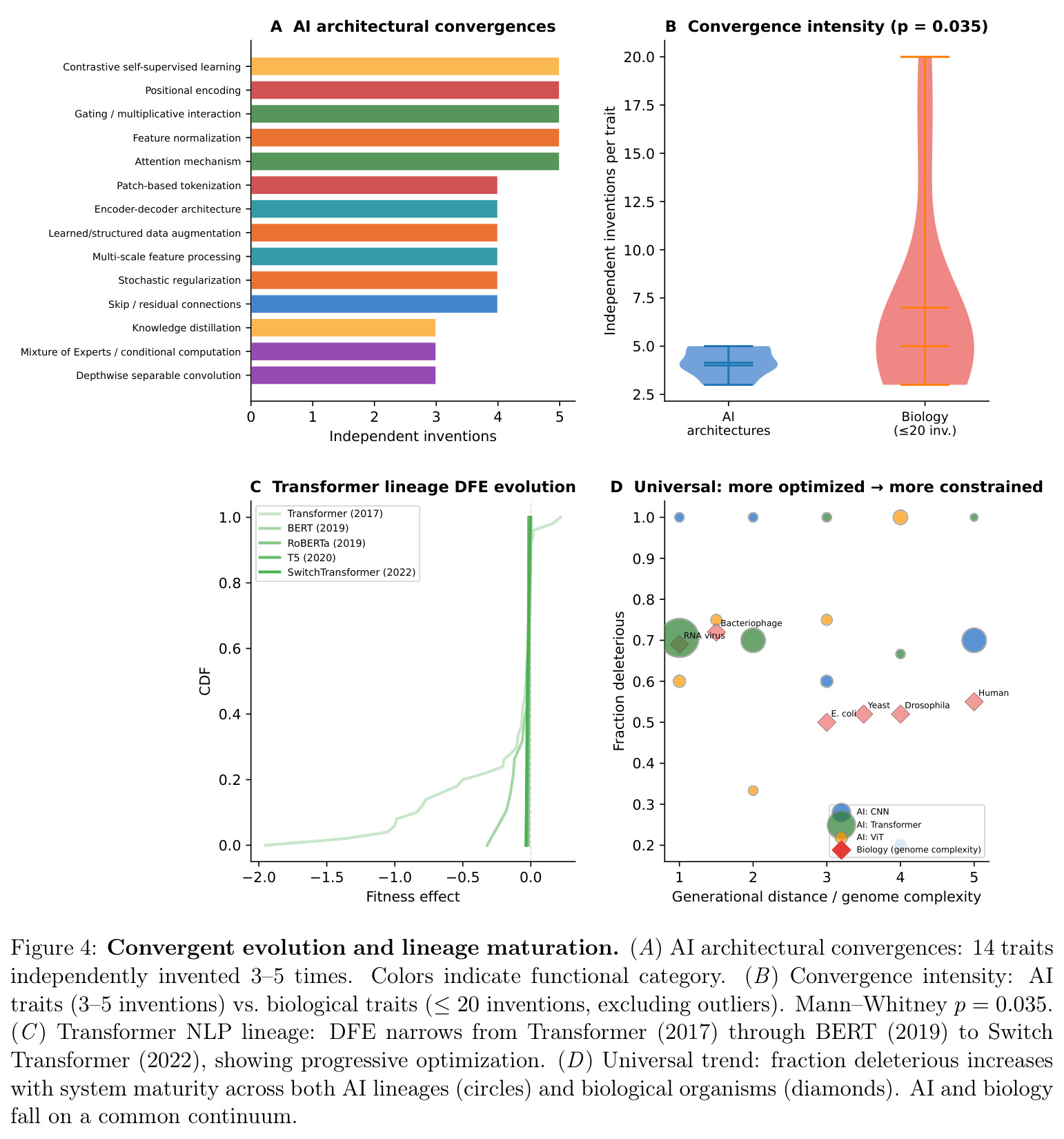
无关谱系在相似选择压力下独立演化出相似结构的现象,如鲨鱼与海豚的流线型体形。本文在 AI 语境下用"不同研究组、不同应用领域独立发明同一架构组件"来类比,并要求至少有 3 次独立起源。
收敛是论证"景观地形有限高适应度解集"的最直接证据,也是把 AI 与生物放到同一证据等级的核心抓手。
研究动机
过去十年 AI 架构从感知机演化到 Transformer、Diffusion、Mamba 等复杂结构,这一过程与生物演化的表面相似性已被多次定性地讨论过——神经架构搜索类比为演化算法、模型缩放定律类比为生物异速生长——但这些比较始终停留在比喻层面。更关键的是,工程化 AI 演化是由人类研究者"带着目的"驱动的,与自然选择的"盲目"机制存在根本差异:当研究者主动尝试可能有益的改动时,其适应度效应分布(DFE)应当呈现何种形态?是否仍服从生物演化中已观测到的重尾、负偏、gamma 拟合的统计规律?这些问题在 2025 年之前没有任何数据集能够定量回答。
本文的目标是本文用 935 次来自 161 篇论文的消融实验(140 次人工整理 + 795 次用 Claude Sonnet 自动抽取)作为 AI 架构演化的"化石记录",系统性地测试三个可证伪假说:H1,AI 架构修改的 DFE 在函数形式与参数上匹配生物 DFE;H2,AI 架构多样化动力学呈现间断平衡与 logistic 饱和;H3,收敛演化的频率与强度与生物可量化可比。目标不仅是描述统计相似性,而是要用 AI 这一加速、可观测的演化系统去检验基础演化理论中悬而未决的问题:演化结构究竟由什么决定?
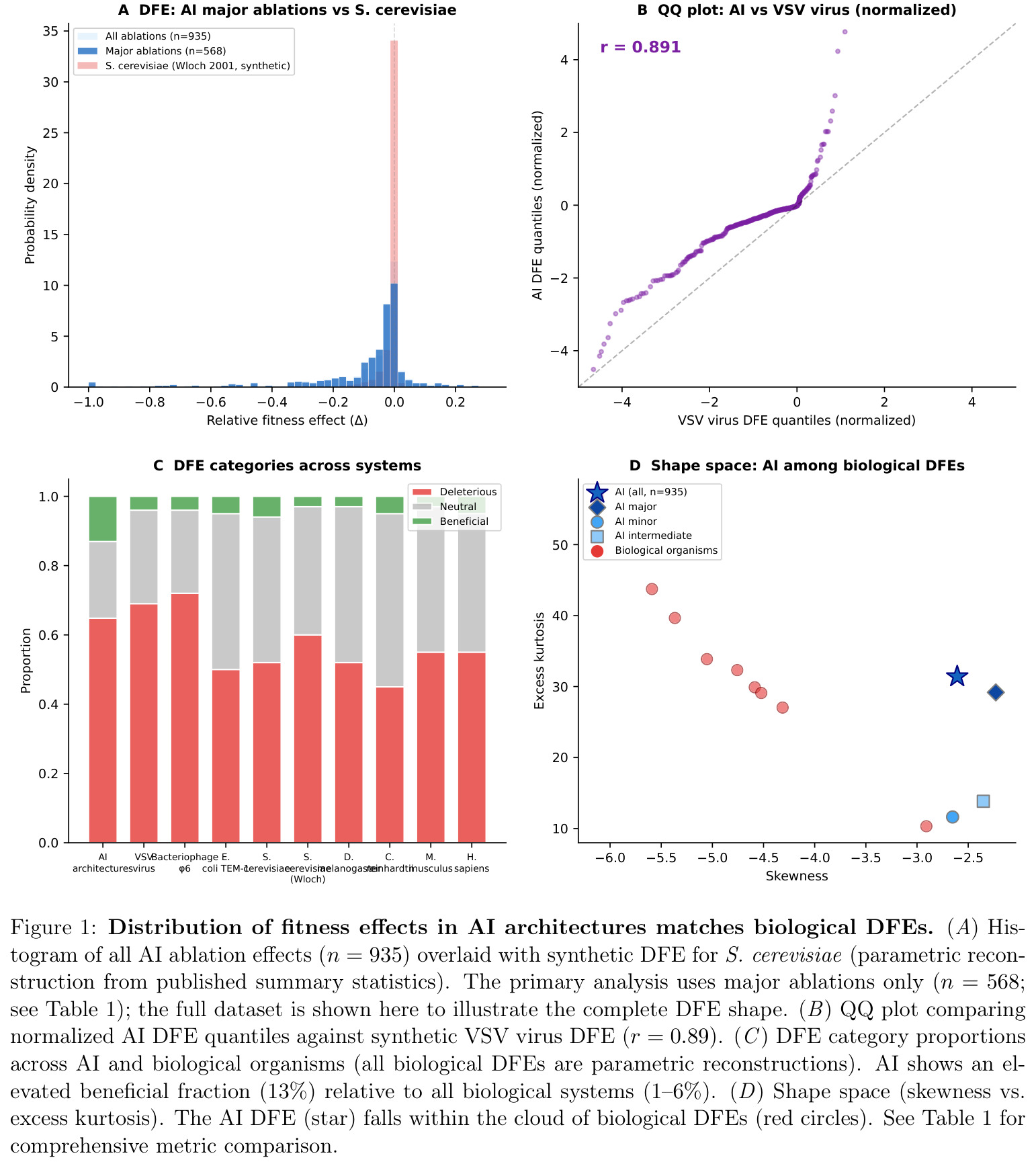
与已有工作不同的是,作者把研究定位为演化理论的"跨基底检验"——若 DFE 形状、多样化曲线、收敛计数三大签名同时在碳基(生物)与硅基(AI)系统上吻合,则说明演化结构由适应度景观拓扑决定,而与变异-选择-遗传的具体物理实现无关。这一切入角度在既有 AI 文献(关心单一架构性能而无视其演化轨迹)和演化生物学文献(缺乏工程化系统的高精度时间序列数据)之间是空白的。本文同时提供方法论贡献——LLM 抽取的 795 条实验比人工整理的 140 条偏差更小,证明在大规模演化数据采集中自动化优于人工。
核心方法
方法论上是一篇大规模经验比较研究,整体思路是把 AI 消融实验"翻译"成生物学的 DFE 测度,再与九种生物(从 RNA 病毒到人类)的已发表 DFE 做统计比较。技术上分四步:数据获取(双轨:人工整理 + LLM 抽取)、相对适应度计算(归一化到完整模型性能)、分布拟合(Student t、gamma、Laplace 通过 AIC 择优)、跨域比较(KS 距离、QQ 相关、欧氏距离、参数估计)。
核心创新在于"基底无关性"的实证范式:把工程化 AI 架构与生物基因组置于同一套演化统计测度下,承认两者搜索机制(盲目 vs 定向)的差异,并把这种差异转化为一个可被检测的预测——AI 的有益突变比例应当系统性高于生物,但分布形态(重尾、负偏、gamma 拟合)应当保留。这与既有"演化类比"研究的关键区别是:作者把景观拓扑与搜索机制这两个变量在统计上分离,只有"分布形状"被保留时,景观假说才能成立。
方法步骤详情
数据获取阶段用 49 个关键词查询从 arXiv 与六个顶会中筛选出 3040 篇候选论文,经关键词打分过滤到 164 篇高置信度候选;每篇论文由 Claude Sonnet 用结构化提示词提取元数据、基线、消融后性能与组件描述,并人工校验 $|\Delta| > 5$ 的 5 条可疑记录后剔除。第二步将消融按规模分为 major($n=568$,完全移除组件)、intermediate($n=154$,替换为更简单替代)与 minor($n=213$,仅调超参),其中 major 是主分析,因其最接近生物学的"基因敲除"。第三步对绝对有害效应 $|\Delta| < 3$ 做最大似然 gamma 拟合,得 AI 的 $\beta = 0.65$(95% CI: 0.59–0.72),并对全分布用 AIC 比较 Student t、Laplace、gamma 三种形式。第四步对生物 DFE 由于个体级数据缺失,采用基于已发表类别比例与 gamma 参数的合成样本做 KS 比较,主要结论仍以不依赖样本重建的参数比较为锚点。
技术新颖性
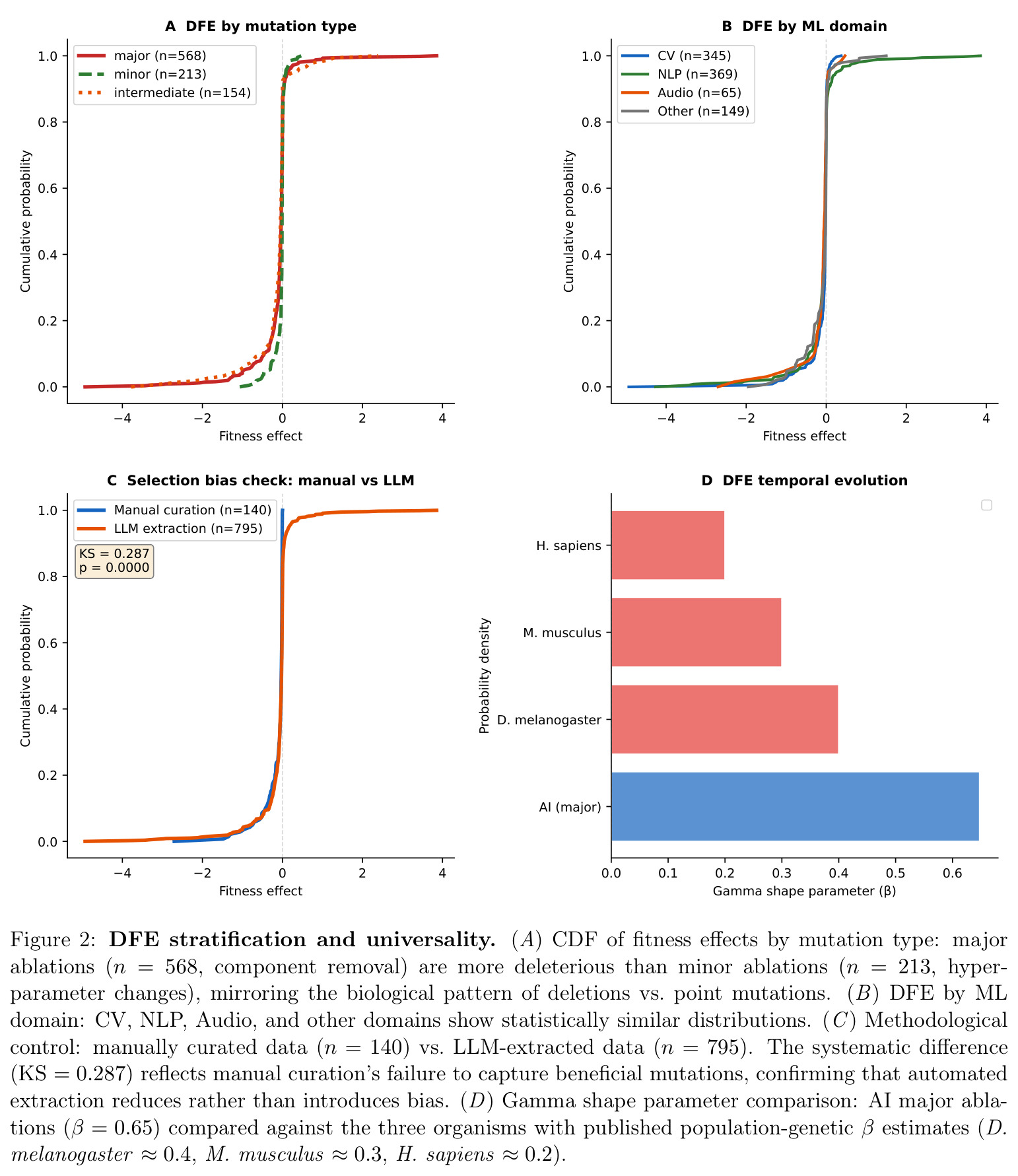
技术新颖性体现在三方面:第一,构建了迄今最大的 AI 消融数据集(935 实验 / 161 篇),数据规模比既有手工整理大近 7 倍;第二,提出"LLM 抽取 + 人工校验"的混合流程,并通过 140 vs 795 的对照(KS=0.287)证明 LLM 数据其实比人工数据偏差更小(人工只采到 0% 有益突变);第三,把 logistic 多样化曲线($R^2=0.994$, $K \approx 142$)与古生物数据库的寒武纪三叶虫、晚白垩世哺乳动物辐射曲线放在归一化时间轴上做形函数比较,这是首次跨如此多数量级时间尺度($10^8$ 年 vs $10^1$ 年)的同形态检验。
实验结果
核心发现分三块。第一,AI 主消融 DFE 为 68.0% 有害(CI: 64.1–71.8%)、19.0% 中性、13.0% 有益,偏度 $-2.23$、峰度 $29.2$,AIC 上 Student t($-428$)显著优于 Laplace 与 gamma;按 KS 距离最接近黑腹果蝇($0.07$)与酿酒酵母($0.09$),按比例欧氏距离最接近噬菌体 φ6($0.11$);$\beta=0.65$ 落在生物范围 $[0.2, 0.6]$ 高端,介于紧凑基因组与简单真核之间。第二,多样化呈 logistic 增长($R^2=0.994$, $K \approx 142$,已饱和 88%),年起源率 CV=$0.53$ 落在寒武纪三叶虫($0.79$)与 K-Pg 后哺乳动物($2.19$)之间,2017 Transformer 与 2021 CLIP+Diffusion 两次峰值构成间断平衡。第三,14 个架构特性被独立发明 $\geq 3$ 次,Transformer 谱系 DFE 随代际变窄,AI 谱系与生物在功能密度轴上沿同一连续分布排列。
查看结构化数据
| 任务 | 指标 | 本文 | 基线 | 提升 |
|---|---|---|---|---|
| DFE 分布形态比较(归一化 KS 距离) | KSnorm(越小越相似) | AI 主消融 DFE 作为基准 | 黑腹果蝇 / 酿酒酵母 / VSV 病毒 / 噬菌体 φ6 | D. melanogaster KSnorm=0.07(最接近),S. cerevisiae KSnorm=0.09,Bacteriophage φ6 dEuclid=0.11(次接近) |
| 有害效应尾部形状 | gamma 形状参数 β | AI β = 0.65(95% CI: 0.59–0.72) | D. melanogaster β≈0.4,M. musculus β≈0.3,H. sapiens β≈0.2 | AI 落在生物范围上端,与 VSV 病毒(log-normal 拟合)不可直接比较但定性一致 |
| 多样化 logistic 拟合 | $R^2$ 与承载量 K | $R^2 = 0.994$,$K \approx 142$,当前 125(88%) | 古生物曲线(Sepkoski 模型) | AI 与寒武纪三叶虫、晚白垩世哺乳动物共享同归一化辐射形态函数 |
| 收敛演化强度 | 独立发明次数(Mann–Whitney p) | 14 个 AI 特性被独立发明 3–5 次 | 生物收敛:3–100+ 次(百万谱系) | 按谱系归一后 AI 收敛强度约 5×10^4 倍于生物,p=0.035(强度分布显著不同) |
| 数据可靠性控制(人工 vs LLM) | KS 距离 + 偏差方向 | LLM 抽取 795 条含 15.3% 有益突变 | 人工整理 140 条 0% 有益 | LLM 偏差更小(KS=0.287,p<0.001 但系统可解释),证明自动化降低而非引入偏差 |
局限与改进
作者明确承认四点局限:(1)数据集基于已发表论文,可能高估成功架构比例;(2)LLM 抽取与人工整理的 KS=0.287 差异虽系统可解释,但仍是潜在偏差源;(3)AI-古生物比较需要在跨越 8 个数量级的时间尺度上做归一化,归一化方法本身引入不确定性;(4)"独立发明"判据弱于生物——ML 研究者跨域阅读论文,存在隐式知识转移,即使最严格标准(无共同作者 + 不同应用域)下注意力也仅有 4 次独立起源,相对生物"数百万独立谱系"的对照是不对称的。此外,观察到的局限还有:n=594 时早期(2014–2018)vs 晚期(2019–2024)架构的 DFE 时序差异显著(p=0.03),n=935 时丢失显著性(p=0.128),表明小样本早期数据贡献了部分信号,结果对样本规模敏感。
独立分析的弱点
独立审视可识别出三处可改进的弱点。第一,biological DFE 比较的"合成样本"路径存在不对称性——KS 距离与 QQ 相关比较的是经验分布(AI) vs 参数重建(生物),这可能低估生物方差,让 KS 数值人为偏小;改进方向是采用共享统计量的贝叶斯后验比较或最优传输距离替代 KS。第二,"不同研究组 + 不同应用域"作为独立发明判据过弱——例如 Transformer 之后的 5 次"注意力机制独立发明"中有相当部分引用了原始 Vaswani 论文,本质是设计扩散而非独立收敛;改进方向是引入知识图谱引用网络度量信息传播距离,仅在引用路径超过一定跳数时才计为独立起源。第三,数据集截止 2024 年,2025 年后的 Mamba-2、SSM、MoE 变体、Diffusion Transformer 等新辐射事件未被纳入,$K \approx 142$ 的承载量估值可能已显著偏离现实,需要用滚动时间窗更新 logistic 拟合。
未来方向
作者已提出四个可证伪预测:起源率随多样度接近 K 而下降、未来辐射事件幅度小于 2017/2021 两次峰值、$\beta$ 将留在 $[0.4, 0.7]$、有益比例因人机协同而上升。这些都是未来 5 年可由持续数据集检验的。此外,论文还隐含三条可延伸方向:(1)把分析扩展到训练超参(学习率、batch size、warmup)层面,构建"超参演化"的独立 DFE;(2)把论文的"高适应度解集有限"思想用于自动架构搜索——既然景观有盆地结构,NAS 算法可先聚类相似 DFE 形状再决定探索方向;(3)将软件突变测试的 DFE 补全为参数化形式,与 AI/生物三方对照,直接回答"重尾 DFE 是任何模块化系统的特征还是演化的特征"。
复现评估
可复现性较好:作者在 GitHub 公开了 935 条消融数据、125 个架构目录、分析脚本与画图代码(https://github.com/mool32/ai-evolution-universal-signatures),并把数据集作为 Supplementary Dataset S1 同行评议发布。生物 DFE 数据来自 Paleobiology Database API(寒武纪三叶虫 5831 taxa、K-Pg 后哺乳 7391 taxa、白垩纪被子 2988 taxa),均为开放数据。复现所需算力极低:935 条样本的 gamma 拟合、KS 检验、logistic 回归均可在普通笔记本上数分钟内完成。潜在复现难度在于 LLM 抽取环节需要 Anthropic API 访问权与结构化提示词工程,单次完整重跑需要数百次 API 调用与数十美元成本;社区若要扩展到 2025 年后的论文则需重新跑抽取流程。
论文图表