基于 RGxEStat 的基因-环境互作显著性与稳定性分析 Significance and Stability Analysis of Gene-Environment Interaction using RGxEStat
RGxEStat:面向育种者的轻量级R交互工具,整合混合效应、AMMI与GGE模型分析G×E互作。
前置知识
基因-环境互作 (G×E)
指同一基因型在不同环境条件下表型表现不一致的现象,表现为基因型排序变化或数值差异,是育种学核心问题之一。
本文所有分析都围绕G×E展开,必须先理解为何同一品种在不同地点/年份产量可能差异巨大。
混合效应模型 (Mixed Effect Model)
线性模型的扩展,兼具固定效应(如品种、地点)和随机效应(如年份、重复),用 REML 或 ML 估计方差分量。
本文显著性分析核心方法,需要 lme4 的 lmer 和 afex 的 mixed 函数配合使用,涉及 Satterthwaite 或 Kenward-Roger 近似。
主成分分析 (PCA) 与奇异值分解 (SVD)
PCA 通过正交变换将高维数据投影到方差最大的低维子空间,数学上等价于对协方差矩阵做特征分解或对数据矩阵做 SVD。
AMMI 和 GGE 模型都用 SVD 分解 G×E 互作矩阵以提取互作主成分 (IPC),是双标图 (biplot) 绘制的基础。
AMMI 与 GGE 双标图 (biplot)
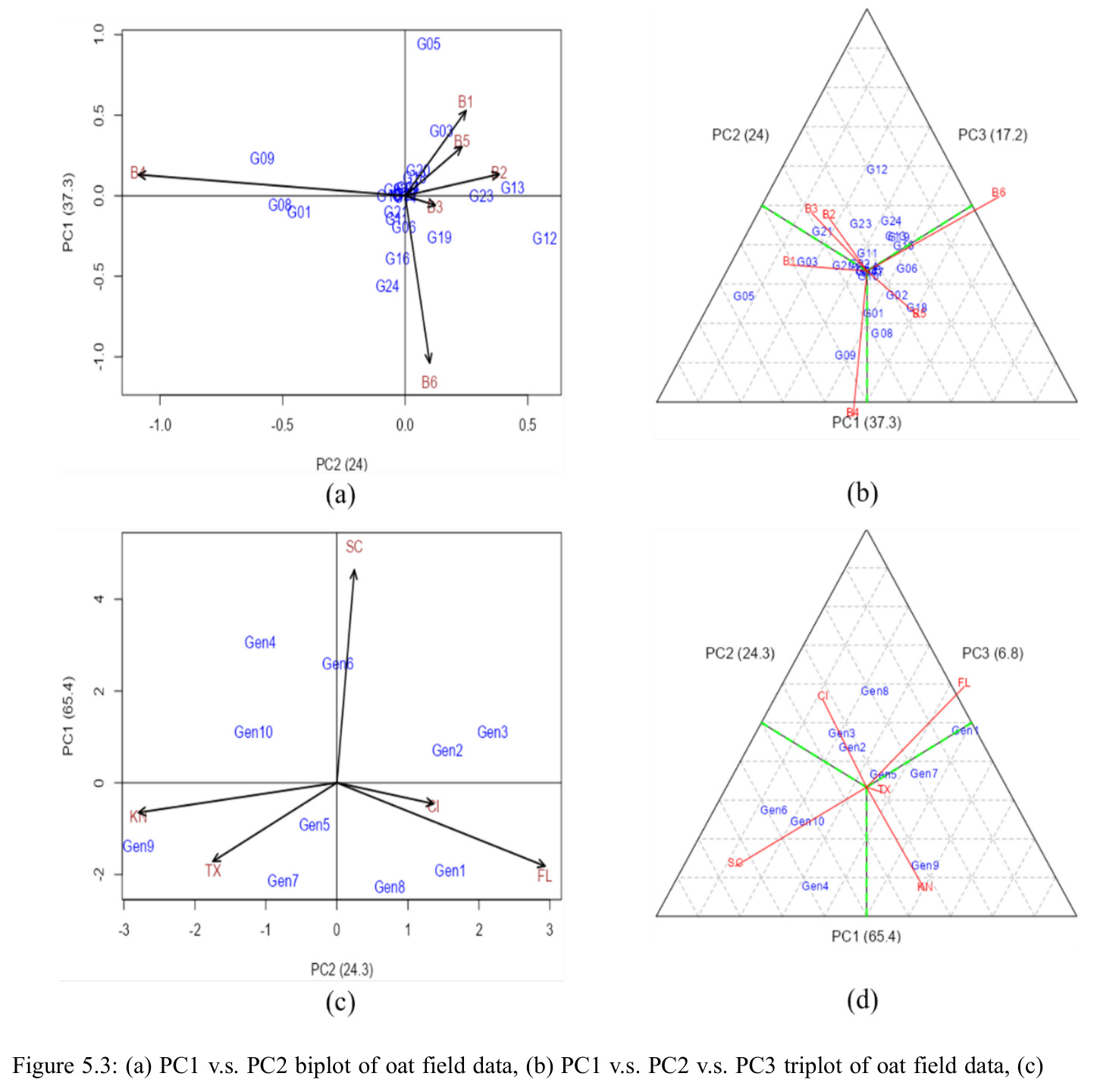
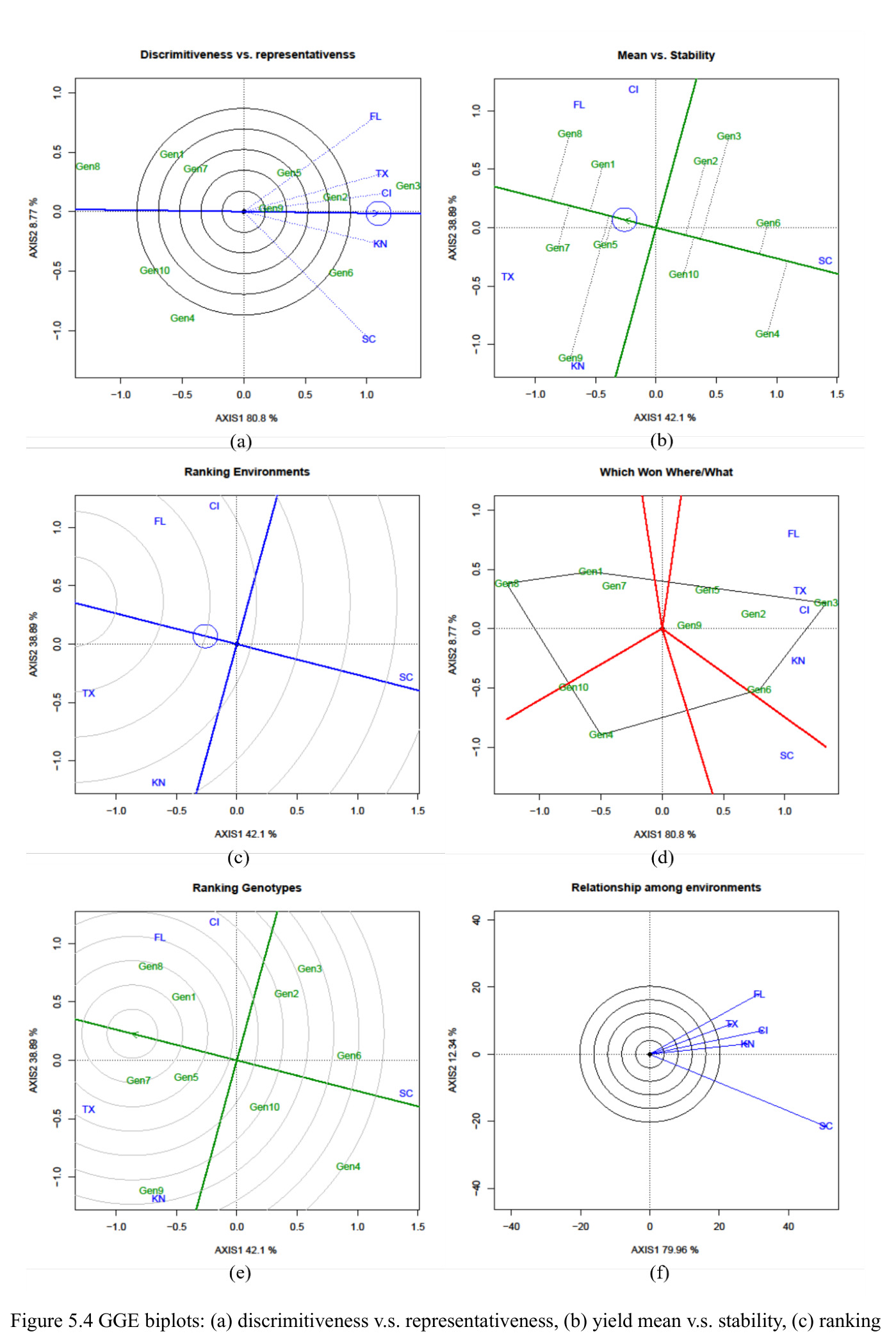
AMMI 拆分为基因主效应+环境主效应+乘性互作;GGE 把基因主效应与互作合并,用 PC1/PC2 同时可视化品种的产量和稳定性。
本文多基因稳定性分析的两大可视化工具,分别用 agricolae 与 GGEBiplotGUI 实现,是判读品种适应性的关键。
假设检验 (t 检验/F 检验/卡方检验)
统计推断基本工具:t 检验用于回归系数是否等于 1,F 检验用于回归偏差是否为零,似然比卡方检验用于嵌套混合模型随机项显著性。
本文单基因稳定性需用 t/F 检验判读斜率和偏差,显著性分析需用 F 检验和卡方检验判读模型各项显著性。
研究动机
在植物育种实践中,育种者需要在多个地点和年份的区域试验中评估品种表现,识别高产且稳定的优良基因型。然而基因型×环境互作 (G×E) 的存在使得同一品种在不同环境下的产量排名和数值可能差异巨大,给选种带来巨大挑战。已有方法存在三大痛点:Piepho 1999 年发布的 SAS 混合模型程序因版本过旧无法在现代服务器上运行;Hussein 等 2000 年的 SASGxESTAB 同样因代码老旧不可用;Dia 等 2016 年发布的新 SAS 代码被作者实测发现部分函数无法实现,且全部方案都要求使用者具备扎实的 SAS 或 R 编程能力,而一线育种者和农艺师通常缺乏这种背景。此外,AMMI 和 GGE 两类多基因稳定性方法的理论之争长期未决,需要一个统一工具让用户无需编程即可完成显著性分析与多种稳定性分析的全流程。
本文的目标是本文的核心目标是开发一个名为 RGxEStat 的轻量级 R 语言交互式软件,将三类核心分析集成到一个图形界面中:一是基于混合效应模型的 G×E 显著性分析(覆盖五种常见情形),二是单基因稳定性分析(计算 Eberhart-Russell 回归斜率/偏差、Shukla 方差、Wricke 生态价、Kang 产量稳定性、Lin-Binns 的 $P_i$、Francis-Kannenberg 的 $CV_i$ 等六类指标),三是多基因稳定性分析(同时支持 AMMI 双标图和 GGE 双标图)。最终目标是把研究者从 SAS/R 代码编写中解放出来,仅通过点击即可完成从数据导入、模型构建、显著性判读到可视化输出的全流程,从而显著缩短育种研究周期。
与已有工作不同的是,本文的独特切入角度在于把统计学的严谨性和工程化的易用性结合起来:方法上覆盖了从单基因到多基因、从显著性到稳定性的完整分析链条;工程上基于 R 的 shiny 或类似交互框架封装了 lme4、afex、agricolae、GGEBiplotGUI 等多个底层 R 包,把零散的方法论整合成一体化流水线。这与 Dia 等 2016 年纯 SAS 论文形成本质区别——后者是方法描述而非工具交付,且其代码被指出存在功能缺陷。本文还贡献了 Forkman-Piepho 参数化自助法选择 AMMI 主成分数 $N$ 这一较新方法的工程实现,这是论文中明确点出的技术细节。
核心方法
RGxEStat 的整体思路是把经典的 G×E 统计方法论封装成模块化流水线。直觉上:用户先上传多年多地点多重复的区域试验数据(包含年份 YR、地点 LC、品种 CLT、重复 RP、目标性状),工具按 YR-LC-RP 嵌套结构自动识别固定/随机效应,调用 lme4 的 lmer 拟合混合效应模型;接着用 afex 的 mixed 函数做 Kenward-Roger 近似下的 F 检验,识别显著的固定效应;对随机效应则通过构造辅助模型做似然比卡方检验;通过显著性后再分两条路径——单基因稳定性按品种做分组回归并计算六类方差统计量,多基因稳定性则对均值矩阵做 SVD 分解并通过 agricolae 与 GGEBiplotGUI 渲染双标图。技术路线上始终遵循"建模→显著性→稳定性"三步走。
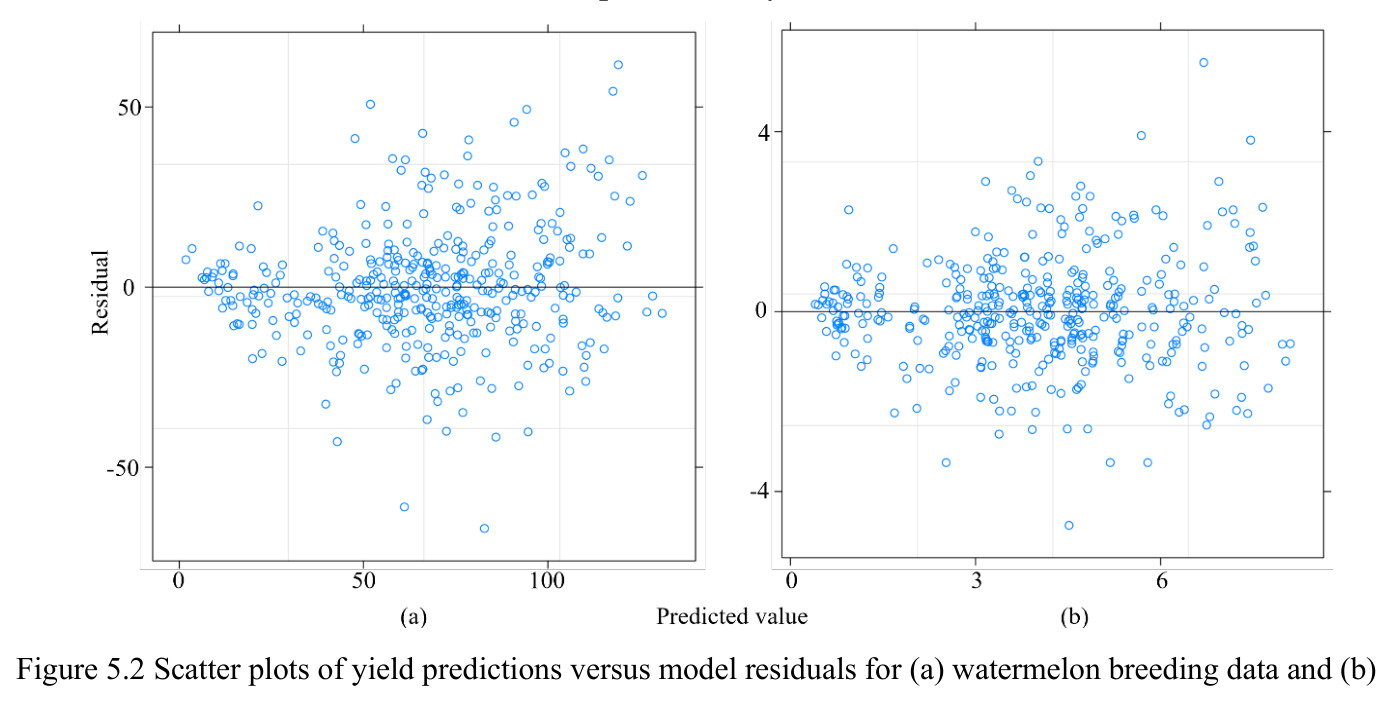
核心创新点在于"工具集成+方法覆盖"两个维度:与已有纯方法论论文(如 Piepho 1999、Hussein 2000、Dia 2016)相比,本文的本质区别是把五类混合效应模型(覆盖 CLT/LC/YR 三因子的全部固定/随机组合)、单基因六类稳定性指标、AMMI 和 GGE 两类多基因模型全部集成在单一 GUI 中,并且明确支持 Forkman-Piepho 参数化自助法选择 IPC 数量 $N$,这是一个相对前沿但工程实现较少的子技术。另一个隐含创新是提供 BLUP 预测的可视化(散点图),让用户直观判断模型拟合质量。
方法步骤详情
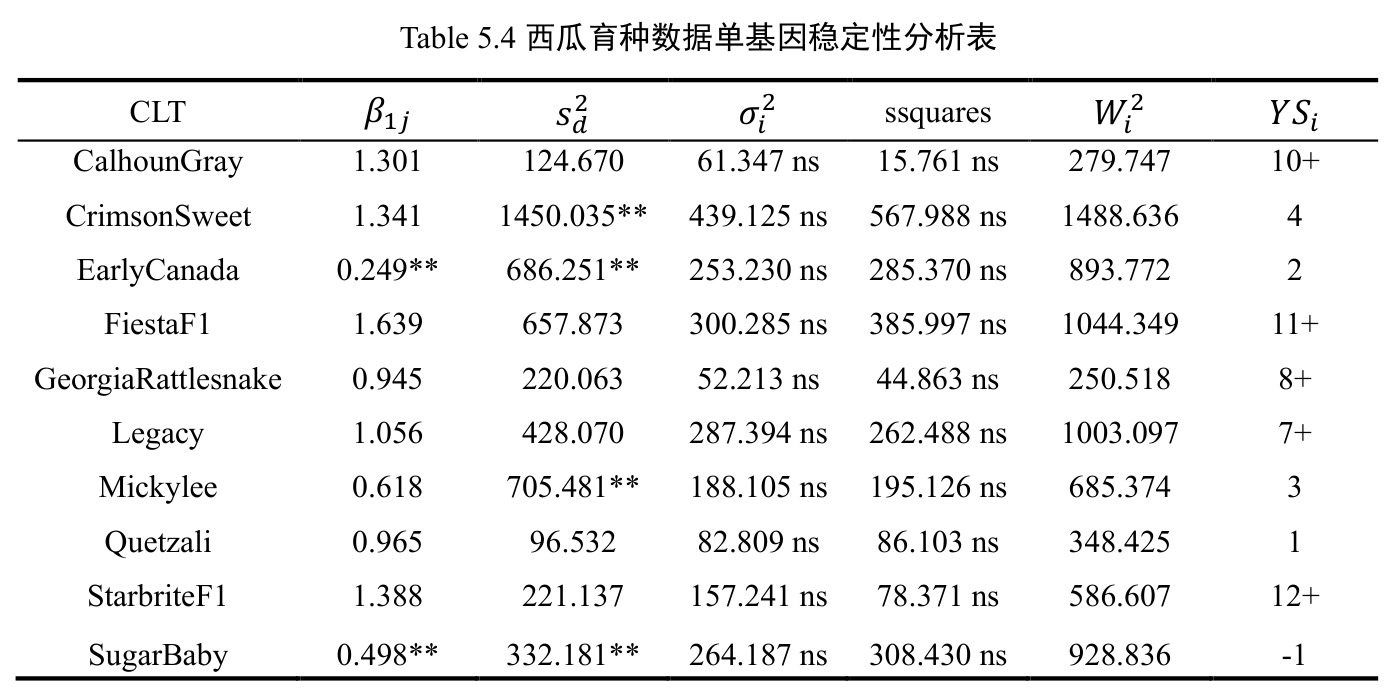
方法分四步。1)混合效应建模:按表 3.1 五种 Case 设定固定/随机属性,用 lme4::lmer 拟合层次模型,固定项经 afex::mixed 的 Kenward-Roger 近似输出 F 检验 p 值。2)随机效应显著性:构造辅助嵌套模型用 stats::anova 做似然比卡方检验。3)单基因稳定性:按品种做分组回归 $Trait_j=\beta_{0j}+\beta_{1j}\cdot ENVTrait+\sum\beta_{Ei}I(ENV=E_i)+\epsilon_j$,对 $\beta_{1j}$ 做 $H_0\!:\beta_{1j}=1$ 的 t 检验、对 $s_{dj}^2$ 做 F 检验;用 agricolae::Stability.par 算 $\sigma_i^2$、$W_i^2$、$YS_i$。4)多基因稳定性:AMMI 用 Forkman-Piepho 自助法选 $N$ 后用 agricolae 画 biplot;GGE 经 SVD 提取 PC1/PC2 后用 GGEBiplotGUI 绘六类双标图。
技术新颖性
技术新颖性主要体现在三个方面:一是首次把 lme4、afex、agricolae、GGEBiplotGUI 等多个 R 包通过交互式界面整合为统一的 G×E 分析流水线,填补了"通才型工具"的空白;二是显式支持 Forkman-Piepho 2014 年提出的 AMMI 主成分数参数化自助选择法($N$ 的选择对结果敏感,这是被点出的关键技术点);三是单基因稳定性模块一次性提供六种指标(含 $W_i^2$、$\sigma_i^2$、$YS_i$、$P_i$、$CV_i$、Eberhart-Russell 回归参数),并指出不同指标在排序上具有相关性(如 $\rho(W_i^2, \sigma_i^2)=1$),给用户对比选择权。局限性是新颖性更多在工程层面,方法学本身都是已有理论的整合而非新方法发明。
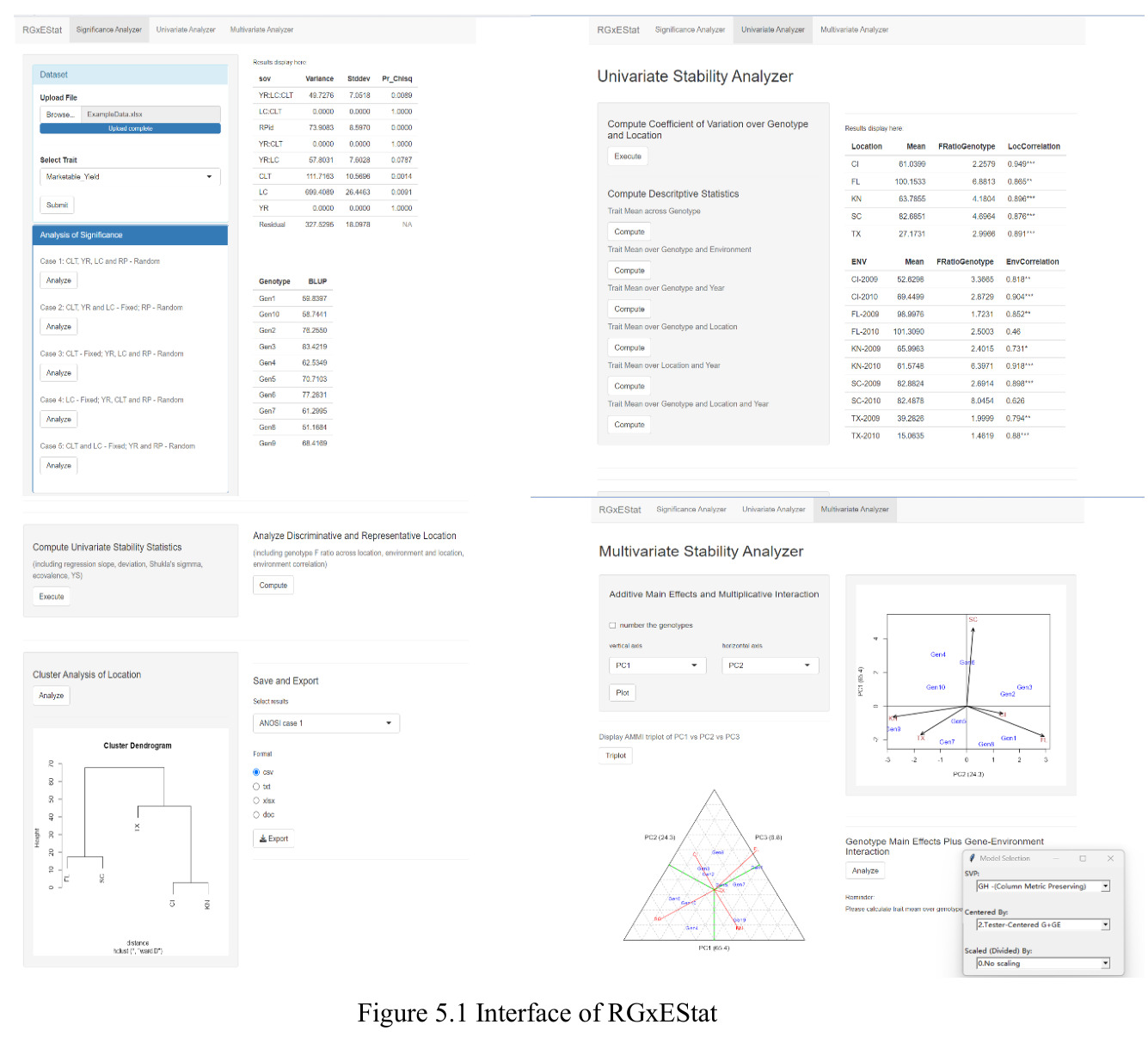
实验结果
西瓜(2 年 10 品种 5 地点 4 重复)Case 1 显示 $\mathrm{YR}\!:\!\mathrm{LC}\!:\!\mathrm{CLT}$ p=0.008、$\mathrm{YR}\!:\!\mathrm{LC}\!:\!\mathrm{RP}$ p<0.001、CLT 方差=111.68(p=0.001)、LC 方差=699.36(p=0.009)显著;$\mathrm{LC}\!:\!\mathrm{CLT}$ 等估计为 0 呈过拟合。燕麦 Case 2 显示 $\mathrm{LC}\!:\!\mathrm{CLT}$ p=0.018、CLT p=0.013、LC p=0.037 三项显著。单基因稳定性识别 CalhounGray、FiestaF1、GeorgiaRattlesnake、Legacy、StarbriteF1 为稳产品种;SugarBaby $\beta_{1j}=0.498$ 显著偏离 1。AMMI 显示 Gen3 在 FL/TX/CL/KN 胜出;GGE 显示 Gen1、Gen5、Gen7 高产稳产;CL、TX 为高区分力试验点。


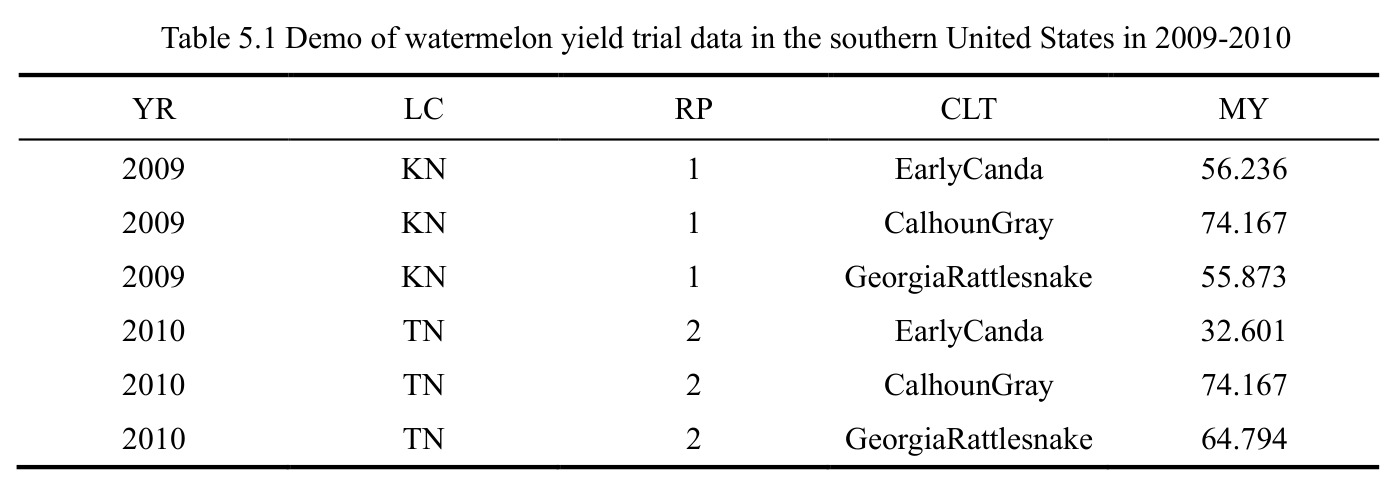
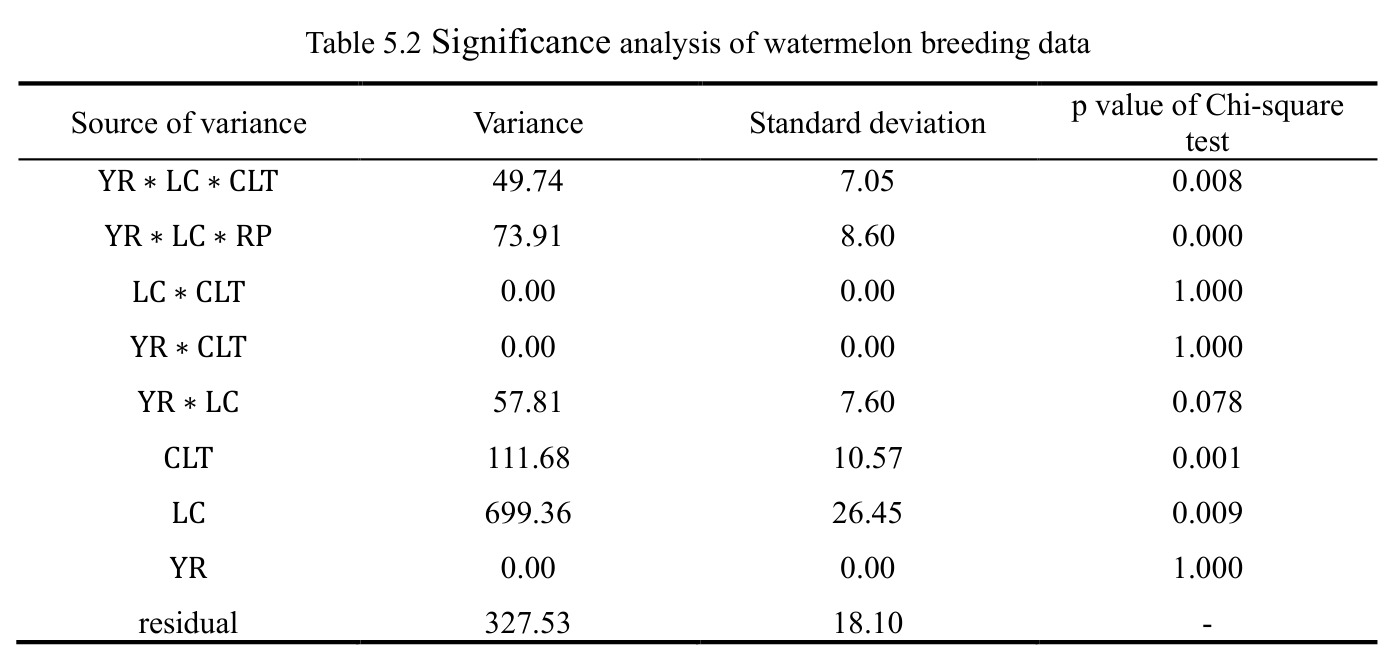
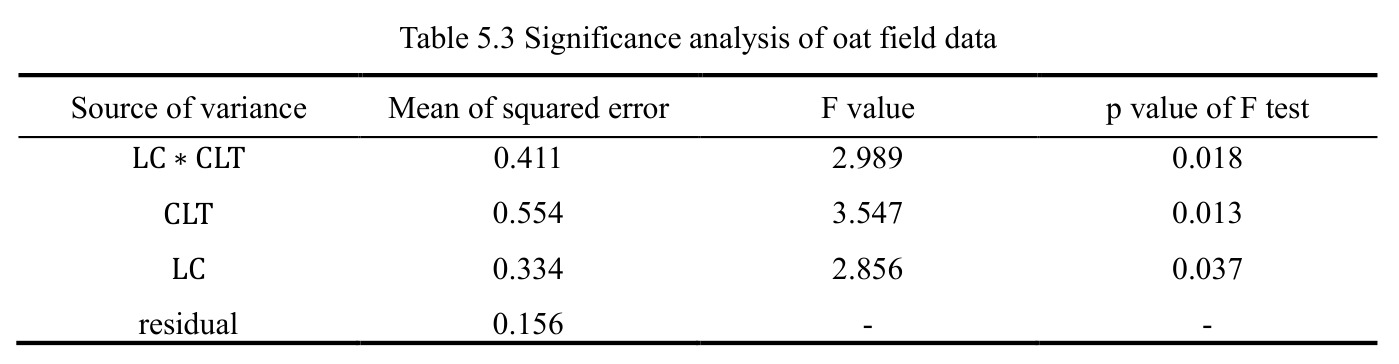
查看结构化数据
| 任务 | 指标 | 本文 | 基线 | 提升 |
|---|---|---|---|---|
| 西瓜数据 YR:LC:CLT 三阶互作显著性 | 似然比卡方 p 值 | p = 0.008 | α = 0.05 阈值 | 在 0.01 水平上拒绝 H0,确认三阶互作显著 |
| 燕麦数据 LC:CLT 互作显著性 | Kenward-Roger F 检验 p 值 | p = 0.018 | α = 0.05 阈值 | p<0.05 拒绝 H0,确认存在 G×E 互作 |
| 西瓜数据品种主效应显著性 | 方差分量(标准差) | 方差 = 111.68,SD = 10.57,p = 0.001 | α = 0.05 阈值 | p<0.01 极显著,品种差异是产量主因之一 |
| SugarBaby 回归斜率偏离 1 的显著性 | t 检验 p 值与 β_1j 估值 | β_1j = 0.498,p < 0.01 | 稳定品种 CalhounGray β_1j = 1.301 不显著 | 判读为低产+对环境变化有较强抗性,识别为不稳定品种 |
局限与改进
作者明确指出的局限有两点:一是方差被估计为零的随机效应在传统 lmer 框架下出现 p 值不可靠的现象(如西瓜数据中 LC:CLT 估计为 0),需要改用 MCMCglmm 的 MCMC 模拟替代;二是对 AMMI 主成分数 $N$ 的选择高度敏感,作者引用 Gauch 2013 年的观点强调 $N$ 选错会导致后续结论偏差,虽引入 Forkman-Piepho 参数化自助法但计算开销随之增加。我观察到的额外局限包括:(1) 论文未在 GUI 上做用户接受度评估,缺少可用性测试数据;(2) 工具仅以 R 包形式发布,对非 R 用户仍存在安装门槛(与"消除 SAS/R 编程需求"的承诺存在张力);(3) 案例数据集规模有限(西瓜仅 10 品种、燕麦仅 24 品种),未测试大规模或超高维场景(如基因组选择中的 G×E);(4) 仅处理单一性状,第六章未来工作也承认无法同时处理多性状 G×E;(5) AMMI 与 GGE 的方法学之争未在工具层面给出选择指南,用户仍需自己判断。
独立分析的弱点
独立分析可见三个具体弱点。第一是用户友好性与技术门槛的张力:作者声称"消除 SAS/R 编程需求",但 RGxEStat 本质仍是 R 包,运行依赖 R 环境和 shiny 服务器,对真正零编程背景的农艺师来说安装配置 shiny 仍是不小门槛;改进方向是提供桌面端可执行文件(如基于 Electron 或 R-Portable 的打包)或托管式 Web 服务。第二是 AMMI 主成分数选择的计算成本:参数化自助法需对残差反复重采样,对大数据集计算开销陡增,且工具未提供并行/降采样选项;改进方向是引入近似 BIC/AIC 准则或 Ledoit-Wolf 形式的收缩估计作为快速替代。第三是多性状分析的缺失:实际育种者往往需要同时优化产量、品质、抗病性等多性状,工具目前只支持单性状;改进方向是引入多变量混合效应模型 (MCMCglmm 的 MVM) 或因子分析模型做联合分析。第四个是显著性诊断的鲁棒性:方差估零导致 p 值不可靠的问题被归咎于 lmer 的 REML 估计,但未给出自动切换到 MCMCglmm 的策略;改进方向是在工具内做方差阈值检测,自动 fallback 到贝叶斯框架。
未来方向
作者在第六章明确提出两大未来方向:一是从统计建模转向深度学习方法,应对高维非线性育种数据,例如用图神经网络建模基因-环境-表型异质网络,或用 Transformer 处理多地点时序数据;二是开展多性状、多基因、多环境 (multi-trait multi-gene multi-environment) 联合分析,因为实际生产中优良性状可能受同一基因控制或彼此独立,简单的逐性状分析会丢失信息。基于 RGxEStat 的工程化成果还可以延伸出三个方向:第一,把当前 GUI 工具进一步与基因组选择 (Genomic Selection) 流水线对接,引入 GBLUP 或 Bayes 系列方法处理 SNP 层面的 G×E;第二,结合迁移学习,把某一环境的 G×E 知识迁移到新环境 (跨域育种),缓解小样本环境数据稀缺问题;第三,把双标图升级为可交互的 Web 双标图(基于 plotly/D3.js),让用户动态筛选品种-环境子集进行探索。
复现评估
复现评估总体良好。代码与数据集均已在 GitHub 开源(https://github.com/mason-ching/RGxEStat),包含两个公开数据集(西瓜为论文作者整理,燕麦来自 R 的 agridat 包),均可在 R 中直接加载。底层依赖的 R 包 lme4、afex、agricolae、GGEBiplotGUI、dplyr、tidyr 都是 CRAN 上的成熟包,安装命令简单。计算资源方面,两组实验数据规模都较小(西瓜 2×5×4×10=400 行、燕麦 6×24=144 行),单核笔记本数分钟即可完成全部分析,不涉及 GPU。复现难度为中等偏低,唯一障碍是 R/shiny 环境的安装配置(约 30 分钟),以及 AMMI 主成分数自助法对计算时间的潜在影响。复现者可严格按论文表 5.2-5.4 的数值核对混合效应模型与稳定性分析结果。
论文图表