基于深度原生结构推理的准确、跨学科且透明的结构-属性理解 Accurate, Interdisciplinary and Transparent Structure-property Understanding with Deep Native Structural Reasoning
SciReasoner通过原生结构推理统一蛋白质、小分子和无机晶体的结构-属性分析
前置知识
结构-属性关系
物质的空间组织、化学连接和周期性排列如何决定其功能、反应性和物理响应。例如,蛋白质的三维折叠决定其生物功能,分子的键合模式决定其化学反应活性,晶体的晶格对称性决定其电子性质。理解这种关系是功能注释、分子设计和材料发现的核心。
论文的核心假设是结构-属性关系无法通过将科学实体视为文本字符串、低维描述符或黑盒输入来完全解决,必须让结构成为推理的原生对象。
自举原生结构推理
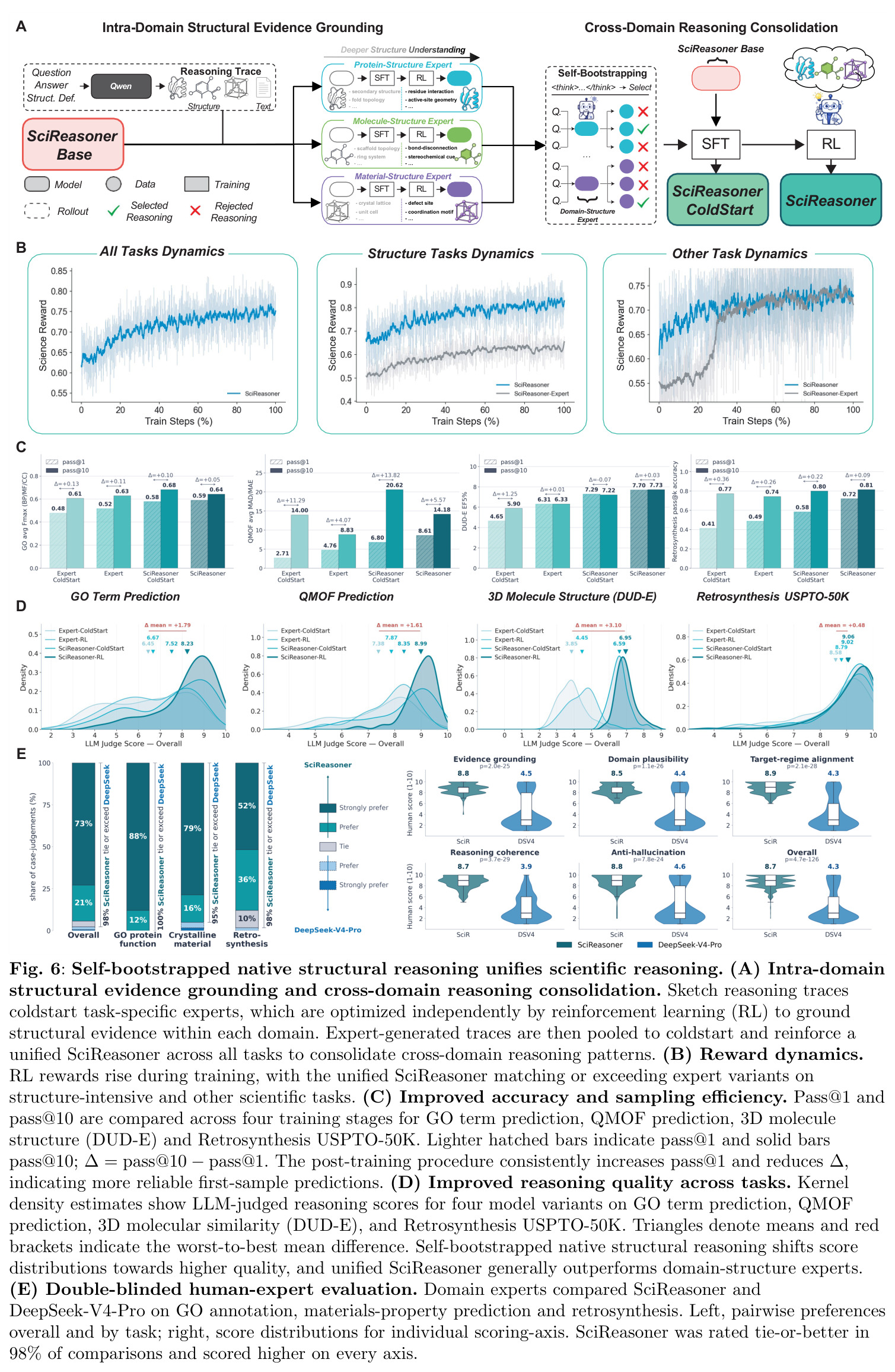
一种后训练框架,包含两个阶段:领域内结构证据基础和跨领域推理整合。第一阶段让领域专家学会如何将结构标记作为推理证据;第二阶段将这些领域本地的证据使用模式整合到统一的能力中,减少标准对齐技术中常见的轨迹崩溃。核心思想是从草图轨迹冷启动专家,然后用专家生成的高质量轨迹训练统一模型。
解决了科学AI中的关键瓶颈:几乎没有能够同时桥接序列、结构和文本的真实推理轨迹,而且异构科学任务的推理模式差异很大,联合训练会导致破坏性干扰。
结构感知词汇表
SciReasoner的离散跨模态投影层,用可学习的嵌入矩阵 $W_v \in \mathbb{R}^{|V_v| \times d_{LLM}}$ 将结构标记序列 $X_v$ 投影到密集连续空间 $H_v = \text{Embedding}(X_v, W_v) \in \mathbb{R}^{L_v \times d_{LLM}}$。这避免了传统多模态框架在推理过程中依赖重型连续编码器的计算瓶颈,同时确保基本物理语义(如局部化学基序、三维几何和晶格对称性)在与语言模型交互之前就作为独特的整体标记被保留。
解决了通用LLM直接应用于科学数据的关键限制:依赖基于文本的子词分词器(如Byte-Pair Encoding)会任意破坏物理拓扑,导致科学语义丢失和训练过程损坏。
Foldseek 3Di标记
一种将蛋白质三维结构编码为离散字母表(如V表示α-螺旋,D表示β-链,L表示环)的方法,与氨基酸序列一一对应。例如,序列段 VVVV 表示稳定的α-螺旋,而 DDLPPVDDP 表示延伸的链/转角区域。这种表示保留了蛋白质的局部结构环境和折叠拓扑,同时与序列保持对齐。
论文中用于结构感知的蛋白质表示,使模型能够引用具体的结构片段(如VVVV映射到规范α-螺旋)作为推理证据,直接将3Di标记子字符串映射到蛋白质的特定空间区域。
SLICES编码
用于晶体的离散编码方法,将晶体结构编码为包含空间群、原子列表和周期性边缘列表的标记序列。例如,+ t a ODD a DOD b OOO c OOO d OOO e OOO o 解码为 Fm-3m 空间群(高对称立方体系),而 0 12 --o 表示钛原子(索引0)与钴原子(索引12)跨越单元细胞边界的周期性连接。这种表示保留了晶体对称性和局部配位环境。
论文中用于晶体结构的原生表示,使模型能够引用具体的对称性和配位模式(如钛-钴周期边缘、钛-镓细胞内边缘)作为稳定性和电子性质预测的推理证据。
ConfSeq
一种将3D分子构象转化为序列兼容表示的方法,结合了分子连接性和内部坐标信息,用 标签包装。它将分子几何特征(如键角、二面角、立体化学)编码为几何感知标记,这些标记来自与自然语言分离的独立词汇表和嵌入表。
论文中用于3D分子表示的关键组件,使SciReasoner能够按结合口袋几何而非2D骨架聚类分子,在DUD-E虚拟筛选中达到5.0% EF = 7.70,超越了之前的最佳结果。
链式思维(CoT)推理
一种推理范式,模型在给出最终答案先生成中间推理步骤。SciReasoner使用格式 $X_a = \langle\text{think}\rangle\n c \n\langle/\text{think}\rangle\n\n y$,其中 $c$ 是潜在推理轨迹,$y$ 是基于 $c$ 的最终答案。关键是,SciReasoner的推理轨迹将自然语言步骤与结构标记(如SMILES片段、3Di片段、SLICES子字符串)交错在一起,使轨迹可以逐个原子检查。
SciReasoner区别于通用聊天模型的关键特征。通用模型只生成自由形式的自然语言解释后跟一个SMILES猜测,而SciReasoner的推理轨迹中的中间步骤本身就是化学结构,可以逐个原子检查,最终反应物组装基于具体的子片段。
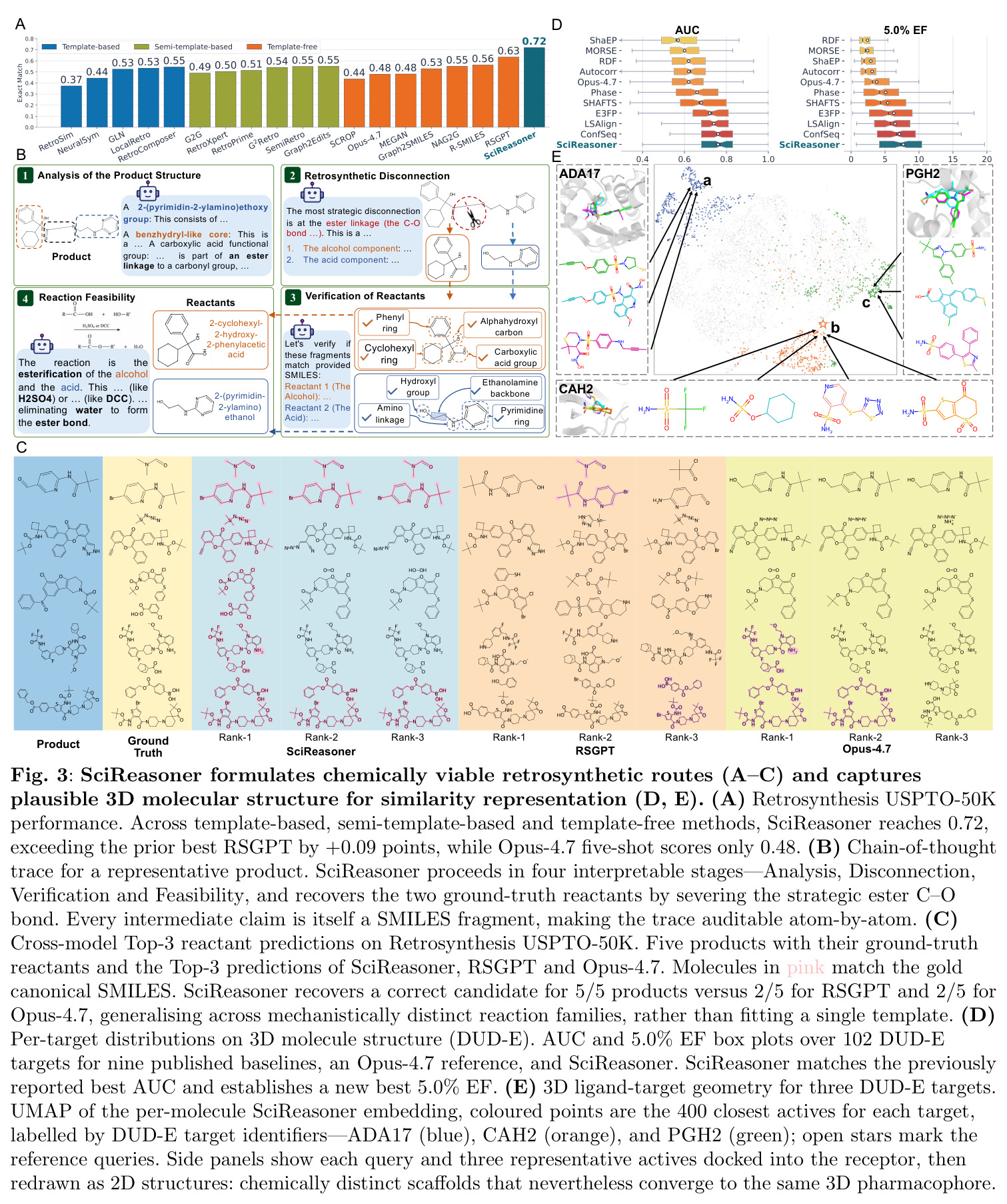
逆合成
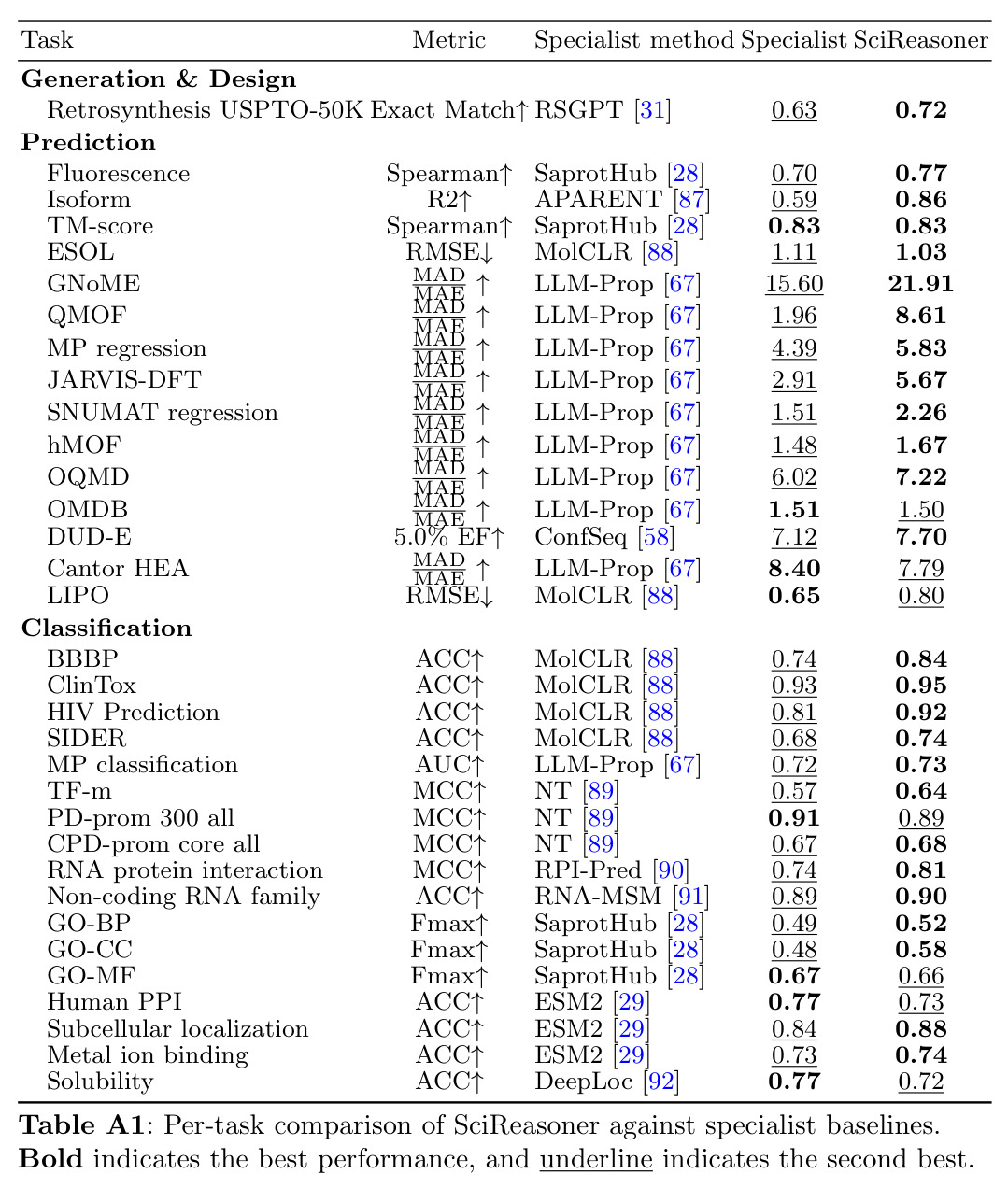
有机合成路线规划的认知骨干,递归地将目标分子分解为商业可用的前体。给定目标SMILES,模型必须输出产生它的反应物SMILES集合。SciReasoner在USPTO-50K基准上将单步准确率从之前的最佳RSGPT的0.63提升到0.72,同时在代表性案例中恢复了5/5的文献反应物(RSGPT为2/5)。
论文中展示SciReasoner化学推理能力的关键任务。不同于模板回忆,SciReasoner通过结构标记的链式思维遵循分析→断键→验证→可行性的工作流程,识别战略性键断开(如酯C-O键)并验证化学上有效的前体。
基因本体论(Gene Ontology, GO)预测
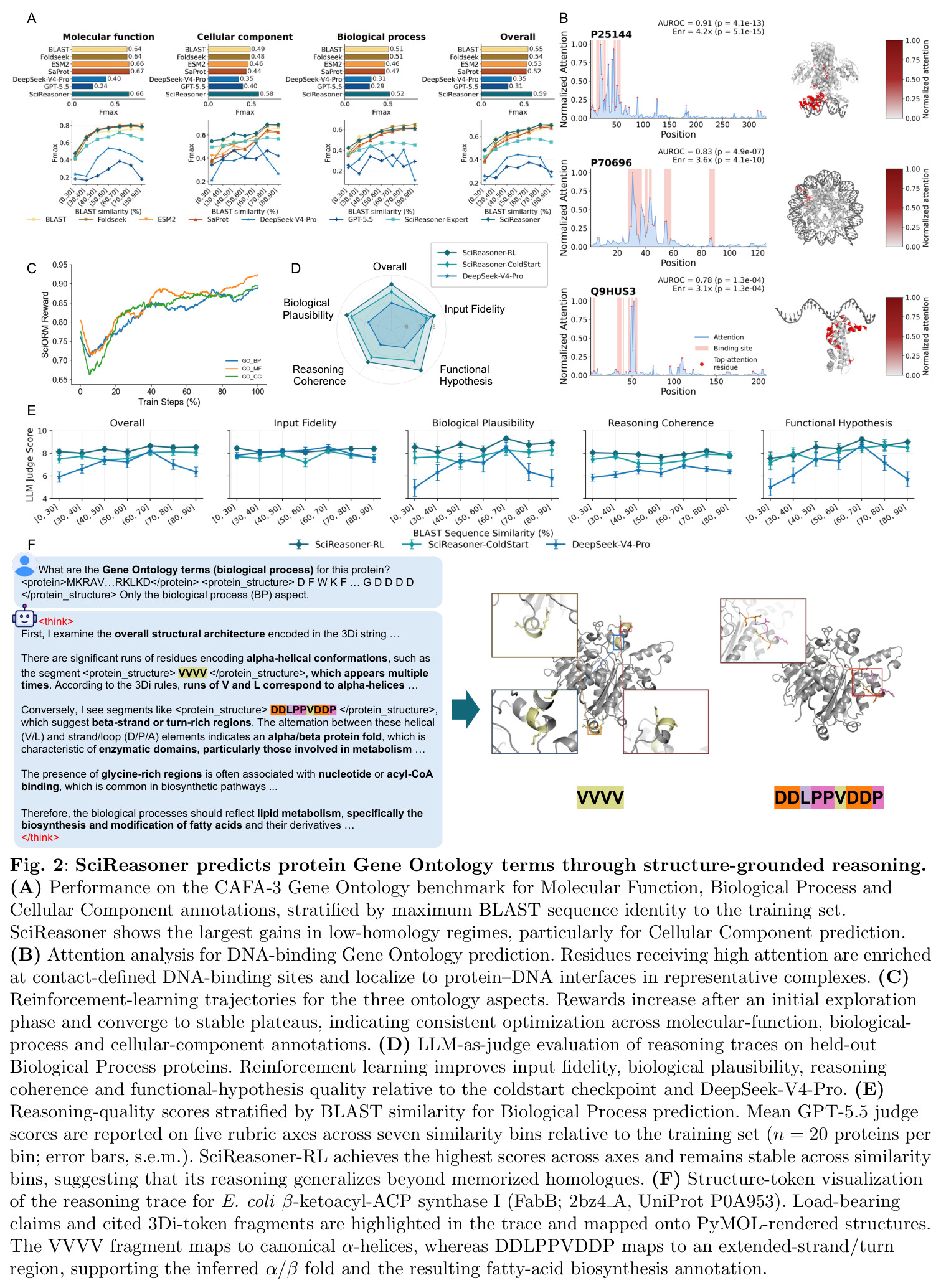
计算生物学的核心任务,将蛋白质序列转化为可解释的功能注释,包括分子功能(MF)、生物过程(BP)和细胞组分(CC)三个方面。论文在DeepFRI-GO基准上评估,特别是在低同源性蛋白质(序列同源性≤30%)上,SciReasoner在CC上的Fmax从0.42提升到0.55,在BP上从0.51提升到0.52。
展示了SciReasoner在生物学推理中的能力,特别是在序列相似性不可靠的低同源性 regime 中的优势。注意力分析显示SciReasoner对DNA结合蛋白质的注意力集中在接触定义的DNA结合残基和蛋白质-DNA界面上。
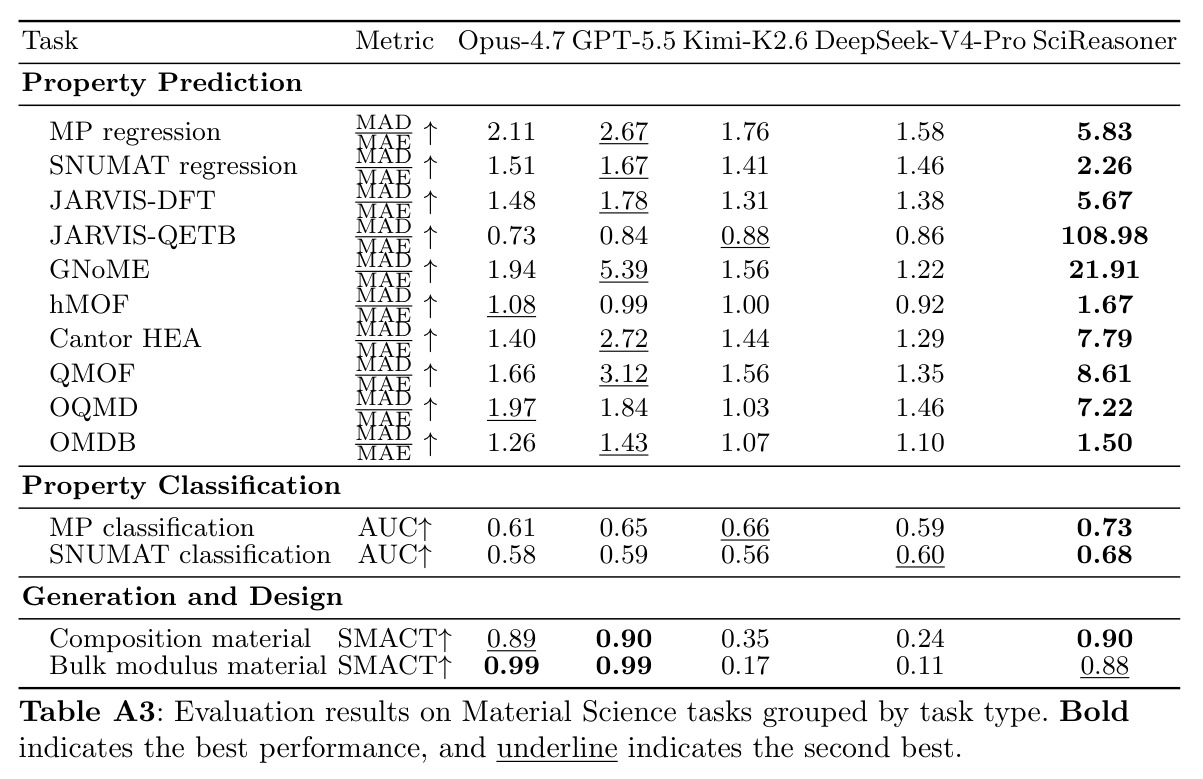
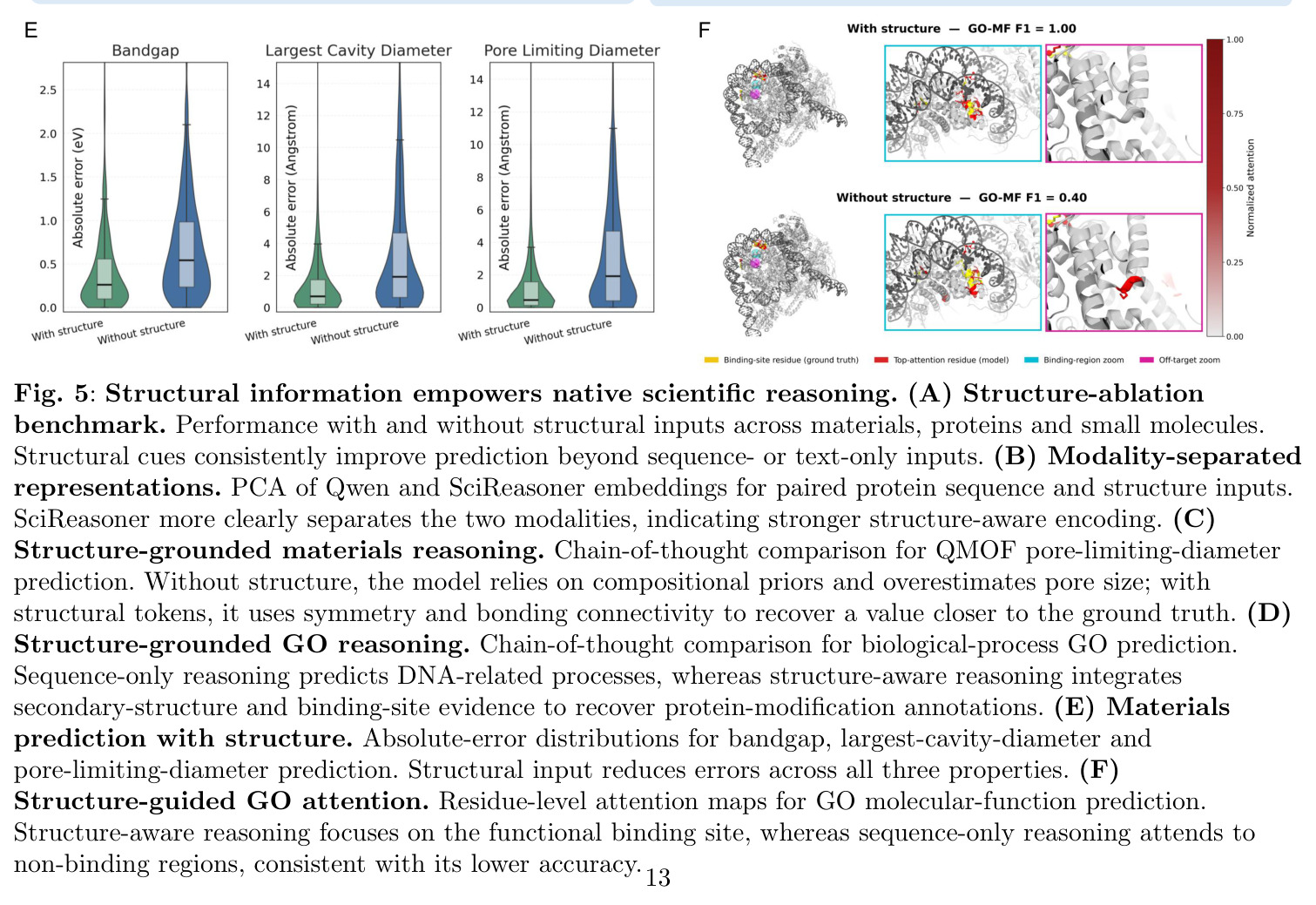
形成能和带隙预测
材料科学中的关键性质预测。形成能反映材料的相对稳定性,带隙决定电子性质(导体、半导体或绝缘体)。论文在Materials Project等数据库上评估,SciReasoner对形成能的预测达到 R² = 0.895,对带隙的预测达到 R² = 0.785,显示了捕获物理上有意义材料-属性信息的能力。
验证了SciReasoner学习结构感知表示的能力。UMAP显示嵌入按化学空间聚类,自发区分不同的结构多晶型,并在每个成分域内形成沿带隙梯度的连续子空间。
虚拟筛选和早期富集因子
药物发现中的关键任务,识别可能结合目标蛋白的化合物。5.0% EF衡量在排名前5%的候选者中活性化合物的富集程度。论文在DUD-E数据集(102个目标)上评估,SciReasoner的5.0% EF达到7.70,超过了之前最好的7.12,AUC达到0.76,与ConfSeq持平。
展示了SciReasoner隐式学习按结合口袋几何而非骨架相似性聚类DUD-E活性化合物的能力。例如,化学上不同的骨架(Morgan指纹相似度仅0.30-0.32)可能收敛到相同的3D药效团。
研究动机
现有科学AI系统在处理结构-属性关系时存在根本性局限。大型语言模型虽然提供了灵活的科学知识接口,但当蛋白质、分子和晶体主要被转化为文本时,结构组织被压缩成字符串或描述,结果解释主要依赖语言关联而非可直接寻址的物理证据。领域特定模型直接编码分子图、蛋白质结构或晶体晶格,但通常优化为任务特定的预测器,输出分数、标签或标量属性,而不暴露其决策背后的中间证据。在低同源性蛋白质功能预测中,BLAST依赖局部序列相似性,ESM2编码进化和序列上下文模式而没有显式结构基础,这些方法在孤儿类蛋白质上表现显著下降。在逆合成中,基于模板的方法受限于已定义反应模板,无模板方法缺乏可检查的中间步骤。在材料科学中,基于文本的模型难以捕获周期性连接和配位环境对电子和热力学行为的决定性影响。
本文的目标是构建一个统一的多模态科学基础模型,能够原生地表示蛋白质、小分子和周期性晶体,并将它们作为可寻址证据单元在生成的推理轨迹中组合、引用和检查。目标是建立一个新的推理范式,使结构成为科学推理的工作基底而非外周输入描述符,让模型不仅预测结构具有什么属性,还开始解释为什么该结构产生特定的功能、反应或材料行为。
与已有工作不同的是,现有方法要么将结构压缩为文本(破坏科学语义),要么将结构作为黑盒输入(无推理可解释性),要么在单一领域内优化(缺乏跨领域能力)。本文的独特切入角度是"原生结构推理":结构标记不是附加到语言的辅助描述符,而是可在生成推理轨迹内组合、引用和检查的可寻址证据单元。通过离散化坐标、拓扑和周期性连接为统一的结构感知词汇表,SciReasoner将结构标记与语言指令集成在单个自回归模型中,能够交错解释文本与具体残基、分子片段、构象线索或晶体描述符。这弥合了两个之前分离的能力:领域原生结构表示和显式科学推理。
核心方法
SciReasoner是一个统一的自回归多模态科学基础模型,由三个主要组件构成:模态特定的离线结构压缩器、结构感知词汇表嵌入层和统一的大语言模型骨干(初始化为Qwen3-14B)。整体训练流程包括三阶段预训练和两阶段后训练。预训练阶段:Stage 1(warm-up)冻结Transformer骨干,仅训练结构感知词汇表、文本嵌入和预测头,建立稳定的结构-语言界面;Stage 2(全参数训练)解冻所有参数,在多样化结构-文本对数据集上训练;Stage 3(退火训练)增加问答(QA)风格数据比例。后训练阶段:首先进行领域内结构证据基础,通过强化学习让领域专家学会将结构标记作为推理证据;然后进行跨领域推理整合,将专家生成的轨迹汇集并训练统一的SciReasoner。这种方法让结构成为推理的原生对象,使模型能够生成可检查的推理轨迹,其中中间声明可以追溯到显式结构证据。
SciReasoner的核心创新是原生结构推理范式,通过统一的结构感知词汇表将三维坐标、分子拓扑和晶体晶格离散化为领域原生标记。与依赖重型连续编码器的传统多模态框架不同,SciReasoner使用离散嵌入查找 $H_v = \text{Embedding}(X_v, W_v)$,优雅地绕过了连续空间编码器的计算瓶颈,同时将结构输入与LLM的原生语义空间无缝对齐。另一个关键创新是自举原生结构推理后训练框架,通过领域内结构证据基础和跨领域推理整合两个阶段,将预训练的结构先验转化为显式科学推理。这个框架解决了科学推理轨迹几乎完全缺失的瓶颈,并减少了标准对齐技术中常见的轨迹崩溃。结构标记与语言指令的交错生成使推理轨迹可以逐个原子检查,实现了预测准确性与可解释科学推理的桥梁。
方法步骤详情
方法包含四个完整步骤:(1)数据构建与处理:蛋白质数据从UniProt和AlphaFoldDB收集,使用Foldseek 3Di编码结构,低置信度区域(pLDDT<70)被屏蔽;小分子数据从ChEMBL和BindingDB收集,使用ConfSeq编码3D构象;材料数据从Materials Project、JARVIS-DFT等数据库收集,使用SLICES编码晶体结构;DNA/RNA数据从RNAcentral和NCBI收集。所有数据通过自然语言模板与文本描述关联。(2)离线结构编码:使用领域特定的离散编码器将原始结构输入编码为结构信息序列:Foldseek用于蛋白质3D结构,SLICES用于晶体,ConfSeq用于3D分子结构。编码后的序列用相应标签包装,如。(3)三阶段预训练:Stage 1(warm-up训练)冻结骨干参数 $\theta_{\text{backbone}}$,仅训练 $\phi = \{\theta_{\text{vocab}}, \theta_{\text{emb}}, \theta_{\text{head}}\}$,在基本结构到文本重建任务上训练,使用独立优化器和学习率调度;Stage 2(全参数训练)解冻所有参数 $\phi = \Theta$,在多样化结构-文本数据上训练,使用共享的Warmup-Stable-Decay (WSD)学习率调度器 $\eta(t)$,包含warmup、stable和decay三个阶段;Stage 3(退火训练)保持全参数优化,增加QA风格数据比例,继续使用WSD调度器。(4)两阶段后训练:首先进行领域内结构证据基础,对每个任务组独立应用DAPO强化学习优化,得到专家模型 ${M_j^{\text{expert}}}_{j=1}^J$;然后进行跨领域推理整合,将专家在各自组提示上滚动的验证通过轨迹 $\tilde{c}$ 替换外部监督 $c$,汇集所有任务并增加非思考数据,从统一冷启动检查点进行单次冷启动和RL训练,得到最终模型 $M_{\text{final}}$。两阶段后训练使用经验求解率过滤和困难中心采样,选择中等难度实例 $\tilde{p}(x) \in (0.125, 0.875)$ 的2000个示例作为每个子任务的RL训练池。
技术新颖性
SciReasoner的技术新颖性体现在三个层面:表示层面,首个将三维坐标、分子拓扑和晶体晶格离散化为领域原生标记的统一词汇表,避免了子词分词器对物理拓扑的任意破坏;架构层面,使用离散嵌入查找替代重型连续编码器,优雅地解决了跨模态对齐的计算瓶颈;训练层面,首次提出自举原生结构推理框架,通过领域内结构证据基础和跨领域推理整合两个阶段,解决了科学推理轨迹缺失和异构任务推理模式冲突的问题。与传统科学AI系统相比,SciReasoner的创新在于让结构标记成为推理轨迹内的可寻址证据单元,而非辅助描述符。这使得残基、分子片段、构象线索和晶体描述符可以在生成推理轨迹中被组合、引用和检查,实现了领域原生结构表示与显式科学推理的桥梁。与通用LLM相比,SciReasoner在结构密集任务上表现显著优势,在低同源性蛋白质GO预测中将CC的Fmax从0.42提升到0.55,在逆合成中将单步准确率从0.63提升到0.72。
实验结果
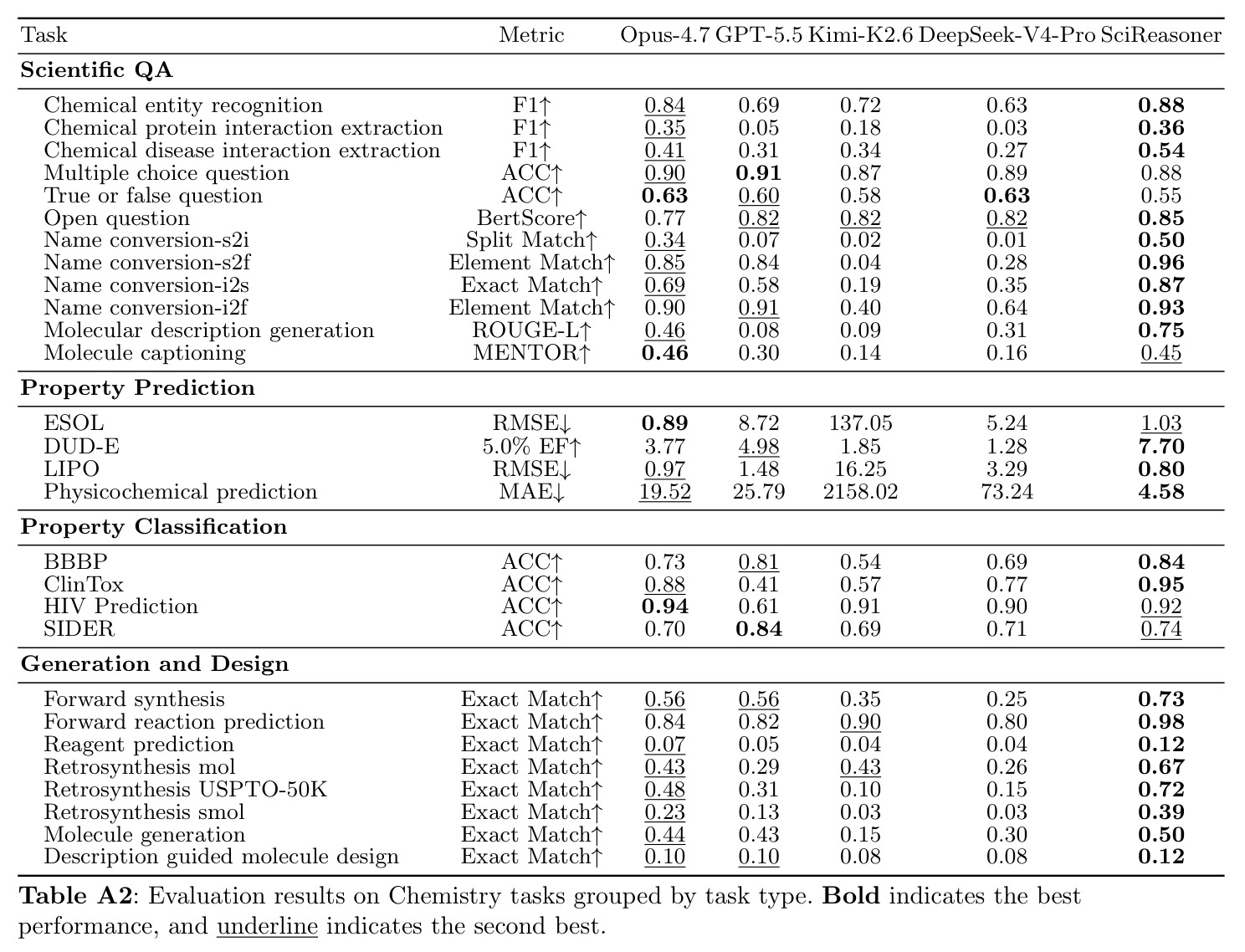
SciReasoner在86个基准测试中的67个任务上达到最先进性能。在蛋白质GO预测的DeepFRI-GO基准上,总体Fmax达到0.59,超越BLAST(0.55)、Foldseek(0.54)、ESM2(0.53)、SaProt(0.52)和DeepSeek-V4-Pro(0.35)。最大的改进在细胞组分(CC)方面,达到0.58 vs BLAST 0.49,特别是在低同源性蛋白质(序列同源性(0, 30%])上,SciReasoner的CC Fmax达到0.55,比BLAST的0.34提高+0.21,比ESM2的0.42提高+0.13。注意力分析显示SciReasoner对DNA结合蛋白质的注意力集中在接触定义的DNA结合残基上,catabolite控制蛋白A达到AUROC=0.91和top-20%富集4.2×,表明功能预测得到物理介导分子相互作用的残基支持。在逆合成USPTO-50K基准上,SciReasoner达到0.72的精确匹配准确率,超越之前的最佳RSGPT(0.63)+0.09点,Opus-4.7五shot仅得分0.48。在5个代表性产品案例中,SciReasoner在Top-3中包含金反应物集5/5,而RSGPT为2/5,Opus-4.7为2/5。在3D分子结构DUD-E虚拟筛选中,SciReasoner达到AUC=0.76和5.0% EF=7.70,匹配之前最好的AUC并建立新的5.0% EF记录。UMAP显示SciReasoner嵌入按结合口袋几何而非骨架相似性聚类活性化合物。在材料科学上,SciReasoner在10个材料属性预测子任务上超越CGCNN,在大多数数值属性上超越LLM-Prop。UMAP显示C、Si和SiC结构清晰分离,形成扩展的子空间区分同成分多晶型。形成能和带隙预测的Parity图显示R²=0.895和R²=0.785,表明捕获了物理上有意义的材料-属性信息。结构消融实验表明,移除结构信息在蛋白质、小分子和材料任务中一致降低性能,证明结构证据是原生科学推理的必要基底而非附加输入通道。双盲专家评估在GO注释、材料属性预测和逆合成三个任务上进行,专家在98%的情况下偏好或平局SciReasoner vs DeepSeek-V4-Pro,其中73%强烈偏好SciReasoner,21%偏好,4%平局。

查看结构化数据
| 任务 | 指标 | 本文 | 基线 | 提升 |
|---|---|---|---|---|
| 蛋白质GO预测(DeepFRI-GO) | Fmax | 0.59 | BLAST 0.55 / ESM2 0.53 / SaProt 0.52 | 总体+0.04 vs BLAST,低同源性CC +0.21 vs BLAST |
| 蛋白质细胞定位 | ACC | 0.88 | ESM2 0.84 | +0.04 |
| RNA蛋白相互作用 | MCC | 0.81 | RPI-Pred 0.74 | +0.07 |
| 逆合成(USPTO-50K) | Exact Match | 0.72 | RSGPT 0.63 / Opus-4.7 0.48 | +0.09 vs RSGPT, +0.24 vs Opus-4.7 |
| 3D分子结构(DUD-E) | 5.0% EF | 7.70 | ConfSeq 7.12 | +0.58 |
| 材料形成能预测 | R² | 0.895 | CGCNN/LLM-Prop未报告 | 高水平相关性, residuals集中在零附近 |
| 材料带隙预测 | R² | 0.785 | CGCNN/LLM-Prop未报告 | 高水平相关性 |
| 虚拟筛选(DUD-E) | AUC | 0.76 | ConfSeq 0.76 | 匹配最佳水平 |
| RNA异构体使用预测 | R² | 0.86 | APARENT 0.59 | +0.27 |
| 蛋白质荧光预测 | Spearman | 0.77 | SaprotHub 0.70 | +0.07 |
局限与改进
作者承认的局限性包括:SciReasoner在某些任务上仍落后于专家模型,如GNoME的MAD_MAE为21.91 vs LLM-Prop 15.60,JARVIS-DFT的MAD_MAE为5.67 vs LLM-Prop 2.91,这可能表明模型在特定材料属性上仍有改进空间。某些分子captioning、化学QA和分类任务上,竞争通用模型保持优势。论文观察到人蛋白质相互作用预测ACC=0.73略低于ESM2 0.77。作者还注意到结构消融实验中移除结构信息在某些任务上性能下降较小,表明可能存在残留的序列或组成级别捷径。从方法角度,自举原生结构推理框架需要两次独立的强化学习运行(领域内和跨领域),增加了计算复杂度。论文的示例主要展示在三个代表性任务上(GO、逆合成、材料),对其他科学领域的适用性尚未充分验证。另外,虽然专家评估显示高偏好,但评估样本量(N=177)相对有限,可能需要更大规模的评估。
独立分析的弱点
SciReasoner在特定材料属性预测上仍存在改进空间,特别是GNoME和JARVIS-DFT等材料数据库上的MAD_MAE指标落后于LLM-Prop。这可能是因为这些属性需要更精细的电子结构计算或实验条件信息,而当前的结构表示可能没有充分捕获这些细微差别。另一个潜在的弱点是,虽然结构消融实验证明了结构证据的重要性,但在某些任务上移除结构信息后性能下降相对较小,这表明模型可能仍在依赖序列或组成级别的快捷方式,特别是在结构信息不充分的情况下。例如,在高同源性蛋白质上,序列相似性可能仍然提供主要信号,掩盖了结构推理的贡献。在计算效率方面,SciReasoner需要两个阶段的强化学习训练(领域内和跨领域),比单一阶段训练更复杂,增加了计算成本和调优难度。另外,当前的方法主要依赖于预训练的结构编码器(Foldseek、ConfSeq、SLICES),这些编码器本身可能存在限制,如对局部环境的敏感性不足或对远程相互作用的编码不完全。最后,虽然论文展示了三个领域的代表性案例,但方法在其他科学领域(如天文学、地球科学)的适用性尚未验证。
未来方向
作者提出的未来研究方向包括扩展到更多科学模态和任务类型,如光谱数据、动力学模拟、量子化学计算和多物理场耦合系统。另一个方向是改进结构表示,特别是对电子结构、激发态和量子效应的编码,这可能需要将密度泛函理论(DFT)计算或波函数信息集成到结构感知词汇表中。训练效率也是一个重要方向,可以探索更高效的自举策略,如并行领域训练或meta-learning方法,减少两阶段强化学习的计算开销。模型解释性可以进一步增强,例如开发结构标记到可视化的自动映射工具,使专家能够更直观地检查推理轨迹中的结构证据。另一个有趣的方向是将SciReasoner与主动学习或实验设计集成,形成从假设生成到实验验证的闭环科学发现系统。可解释性方面,可以开发结构敏感注意力可视化工具,让专家看到哪些结构特征驱动了模型的预测。最后,大规模的专家评估和用户研究可以更好地理解SciReasoner在实际科学工作流程中的效用,并识别需要改进的具体领域。
复现评估
论文提供了详细的实验设置、数据来源和训练参数。代码、模型和数据集的可用性信息可以在项目网站SciReasoner.github.io上找到。预训练数据集来自公开数据库(UniProt、AlphaFoldDB、ChEMBL、BindingDB、Materials Project、JARVIS-DFT、SNUMAT、hMOF、QMOF、OQMD、OMDB、GNoME、Cantor HEA、RNAcentral、NCBI等),但具体的数据处理和过滤步骤的代码开源情况未明确说明。模型架构基于Qwen3-14B,这是一个公开可用的基础模型。训练使用了三阶段预训练和两阶段后训练,每个阶段的学习率调度、数据比例和优化器配置都有描述,但具体的超参数(如warmup步数、stable步数、decay函数形式)在论文中未完全披露。算力要求方面,由于使用了14B参数的模型,预训练和后训练需要大量GPU资源,论文未报告具体的训练时间和硬件配置。评估基准都是公开数据集,评估指标的定义清晰,这使得结果可以复现。然而,某些实验设置(如RL训练中的采样温度T的自适应调整、困难中心采样的具体实现细节)的代码开源情况未知,可能影响复现。总体而言,论文提供了足够的方法细节,但完整的复现可能需要访问源代码和训练脚本。
论文图表
图1展示了SciReasoner的整体架构和训练流程。子图A显示了结构感知科学推理接口,SciReasoner将异构科学实体(蛋白质/DNA/RNA序列、小分子、蛋白质结构、晶体结构、3D分子)编码为结构感知标记,与语言指令结合后由LLM骨干处理。子图B展示了结构感知标记构建,使用领域特定编码器(ConfSeq用于分子、Foldseek用于蛋白质、SLICES用于晶体)将科学对象转换为保留化学和物理意义的结构化文本表示。子图C展示了分词器压缩和语义保持,与Qwen分词器相比,SciReasoner分词器减少了分子输入的token长度,同时保持化学上有意义的子结构。子图D展示了持续预训练和后训练流水线,包括三阶段预训练(warm-up、全参数、退火)和两阶段后训练(领域内结构证据基础、跨领域推理整合)。子图E展示了基准性能,SciReasoner在多样化科学QA、生物学、化学、材料科学、生成/设计基准上超越通用语言模型和领域特定基线。
Figure 1是理解SciReasoner整体架构、设计原理和训练流程的核心图,展示了从结构编码到训练策略的完整管线,对于理解论文的技术路线至关重要。
表A4展示了生物学任务按任务类型分组的评估结果。在科学QA类别中,SciReasoner在功能上达到ROUGE-L 0.80远超所有基线(最高Opus-4.7仅0.30),在一般功能上达到ROUGE-L 0.77超越Opus-4.7 0.52;在属性预测类别中,SciReasoner在荧光上达到Spearman 0.77超越Opus-4.7 0.44,在稳定性上达到Spearman 0.61超越Opus-4.7 0.36,在增强子活性上达到HK-PCC 0.64超越Opus-4.7 0.07,在异构体上达到R²=0.86远超所有基线(最高Opus-4.7仅0.09),在TM-score上达到Spearman 0.83远超所有基线(最高Opus-4.7仅0.15);在属性分类类别中,SciReasoner在溶解性上达到ACC 0.72超越Opus-4.7 0.58,在RNA蛋白相互作用上达到MCC 0.81远超所有基线(最高DeepSeek-V4-Pro仅-0.13),在GO-BP上达到Fmax 0.52超越Opus-4.7 0.43,在GO-CC上达到Fmax 0.58超越Opus-4.7 0.33,在GO-MF上达到Fmax 0.66超越Opus-4.7 0.47,在亚细胞定位上达到ACC 0.88远超所有基线(最高GPT-5.5仅0.65);在生成与设计类别中,SciReasoner在功能引导蛋白设计上达到Normalized SW 0.94超越Opus-4.7 0.74,在催化活性上达到ROUGE-L 0.70超越Opus-4.7 0.60。
Table A4是理解SciReasoner生物学任务全面性能的关键表格,展示了从QA到预测到分类到生成与设计的各类生物学任务上SciReasoner的显著优势,特别是在GO预测、蛋白质功能和生成任务上的出色表现。