几何相变赋予海马体极限记忆容量 Geometric Phase Transition Enables Extreme Hippocampal Memory Capacity
储食鸟类海马通过拓扑相变形成晶体式几何编码,容量提升超百倍。
前置知识
表征差异矩阵 (RDM)
RDM 是 $N \times N$ 矩阵,元素 $(i,j)$ 记录群体对刺激 $i$ 与 $j$ 的响应差异(如余弦距离),用于刻画神经群体的几何结构。
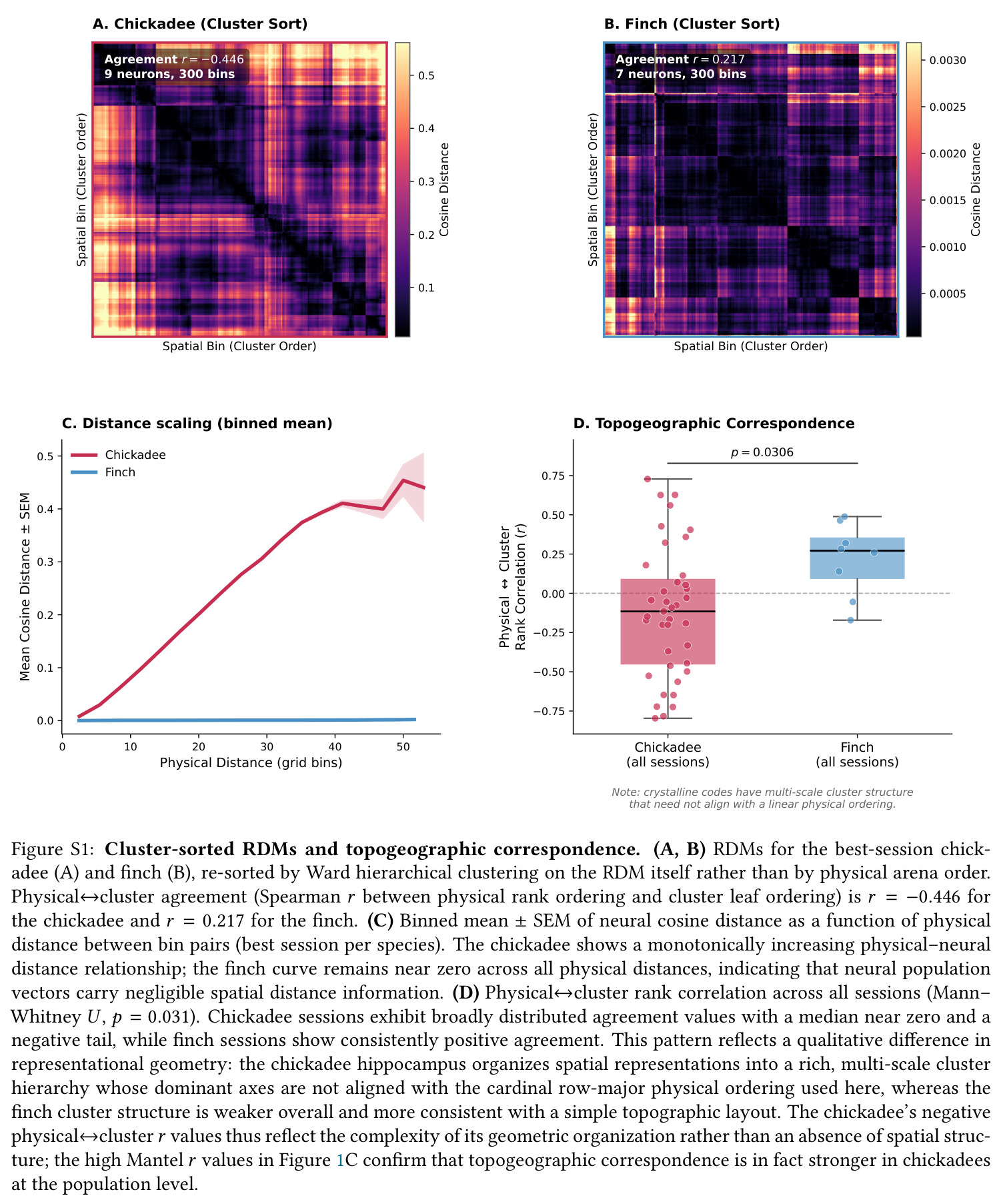
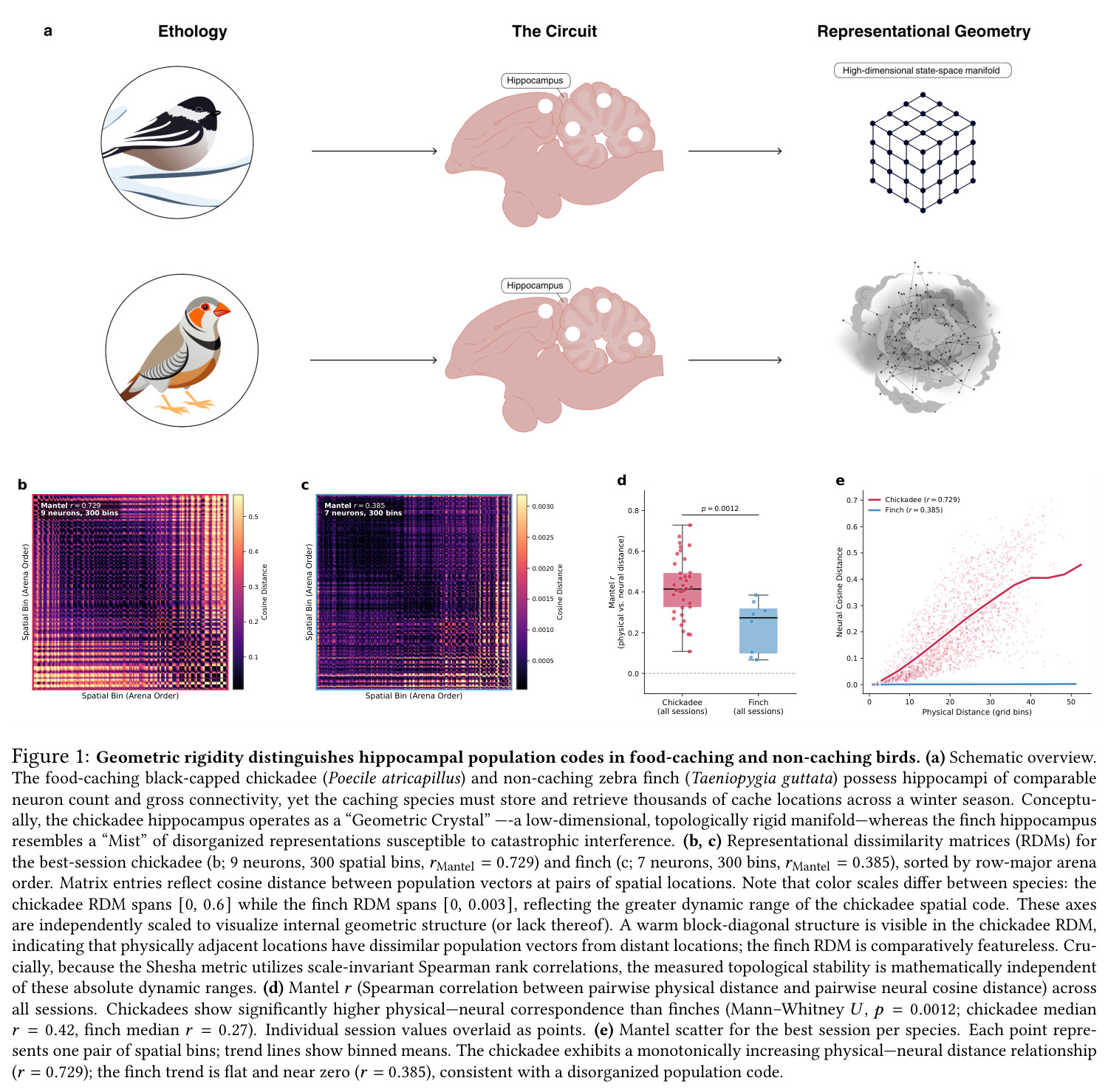
本文核心证据 chickadee vs finch 的 RDM 块对角结构差异直接来源于此概念。
Mantel 检验
通过置换检验比较两个距离矩阵(如物理距离矩阵与神经 RDM)的 Spearman 相关性,评估神经编码是否反映物理拓扑。
chickadee 的 $r_{\text{Mantel}}=0.420$ vs finch 的 $0.230$ 是本文第一关键统计量。
主角度 (Principal Angles)
用于度量两个线性子空间正交程度的几何量,角度越接近 $90°$ 代表子空间越独立,越接近 $0°$ 代表越重合。
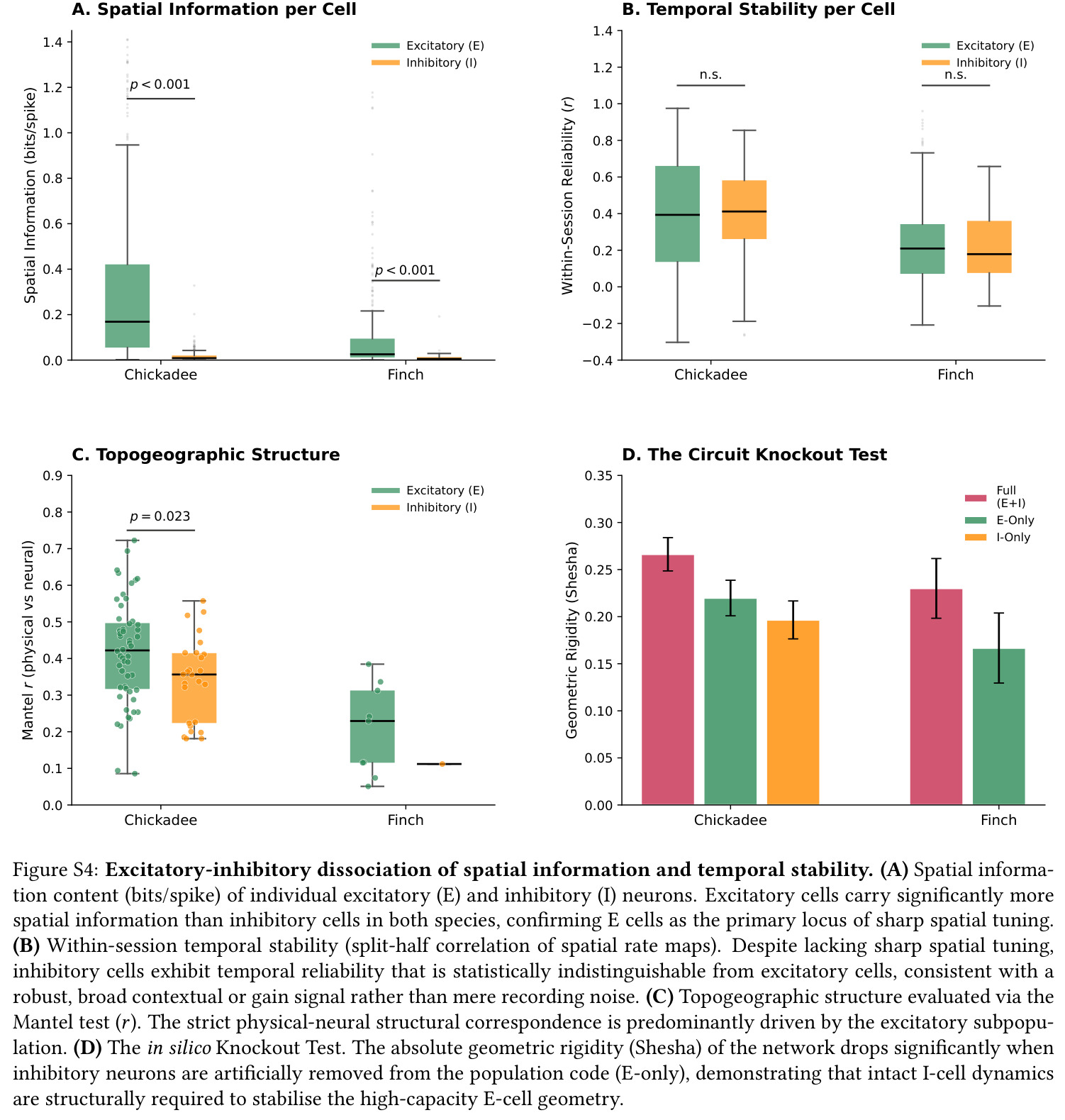
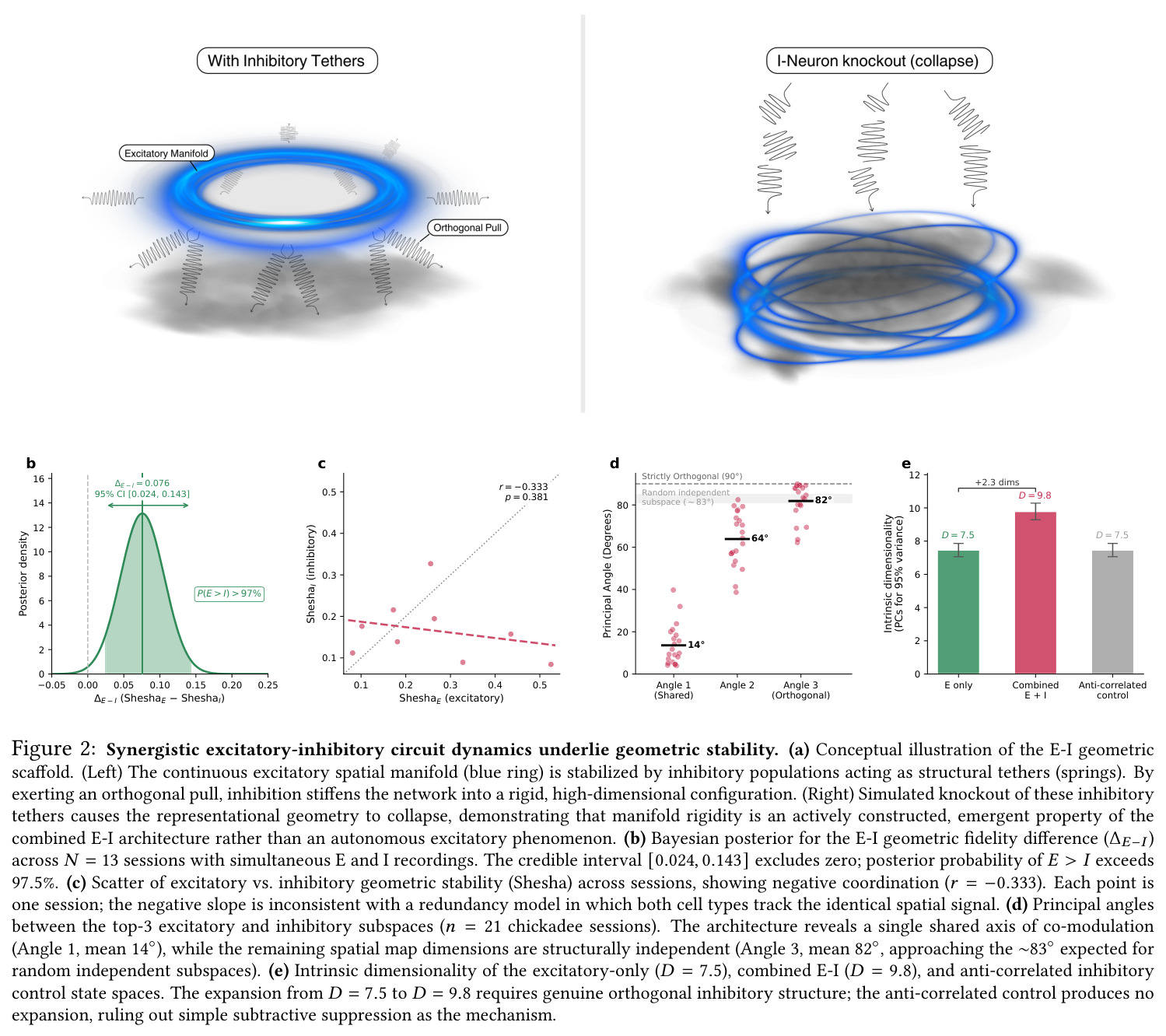
本文兴奋/抑制群在 $14°, 64°, 82°$ 的'一共享、两正交'模式依赖此概念。
分裂半可靠性 (Split-half reliability)
将神经元随机均分为两个子集,分别独立计算编码指标后再求相关,用以衡量该指标对采样噪声与子集选择的稳健性,Shesha 即是该思路在 RDM 上的应用。
Shesha 几何稳定性指标本质上是分裂半 RDM 相关,是核心方法学。
灾难性干扰 (Catastrophic interference)
神经网络在学习新记忆时旧记忆被灾难性覆盖的现象,是生物学记忆容量受限的根本原因,也是储食鸟类必须解决的核心计算瓶颈。
本文论证几何刚性是抵御灾难性干扰的'机制性药物',容量差 >100 倍即源于此。
本征维度 (Intrinsic dimensionality)
群体响应有效占据的独立方向数,常用参与比 $D = (\sum_i \lambda_i)^2 / \sum_i \lambda_i^2$ 度量。
抑制群使 $D$ 从 $7.5$ 升至 $9.8$ 是 E-I 协同效应的关键量化。
研究动机
生物记忆系统面临一个根本性的尺度悖论:储食的黑顶山雀(black-capped chickadee)能在整个冬季记住数千个食物埋藏位置,而非储食的斑胸草雀(zebra finch)则只能维持极有限的空间记忆。两者海马神经元数量与基础连接性相当,经典框架(如 Marr 1971、Hopfield 1982、Treves-Rolls 1994)只能解释容量随硬件规模线性或多项式增长,无法解释为何相同的生物硬件能产生数量级差异的记忆能力,更无法解释随记忆负载上升时如何避免灾难性干扰(catastrophic interference)。
本文的目标是本文旨在系统揭示储食鸟类海马获得超常记忆容量的神经计算原理:通过对比储食黑顶山雀(39 sessions, 9 birds)与非储食斑胸草雀(8 sessions, 9 birds)在开旷场(open-field)觅食任务中的群体编码,量化两物种在表征几何(Mantel 与 Shesha 指标)、兴奋-抑制回路动力学(主角度与本征维度)、以及合成网络容量曲线上的差异,并寻找一个能跨越经验数据与计算模型一致的几何相变机制作为统合解释。
与已有工作不同的是,与既有研究多聚焦于单神经元位置野(place field)精度或神经元数量调动的传统思路不同,本文跳出离散神经分配模型(Valiant 的 Stable Memory Allocator 框架),转向以群体流形的拓扑刚性(topological rigidity)作为认知极限的组织原则——核心主张是:海马进化不是靠堆叠神经元的'算术增殖',而是靠工程化群体编码本身的几何结构来获得极限容量。这一'几何相变'视角既能与 E-I 协同回路机制对接,又能解释山雀与草雀在硬件相当下数量级的行为差异,是当前理论神经科学的关键空白。
核心方法
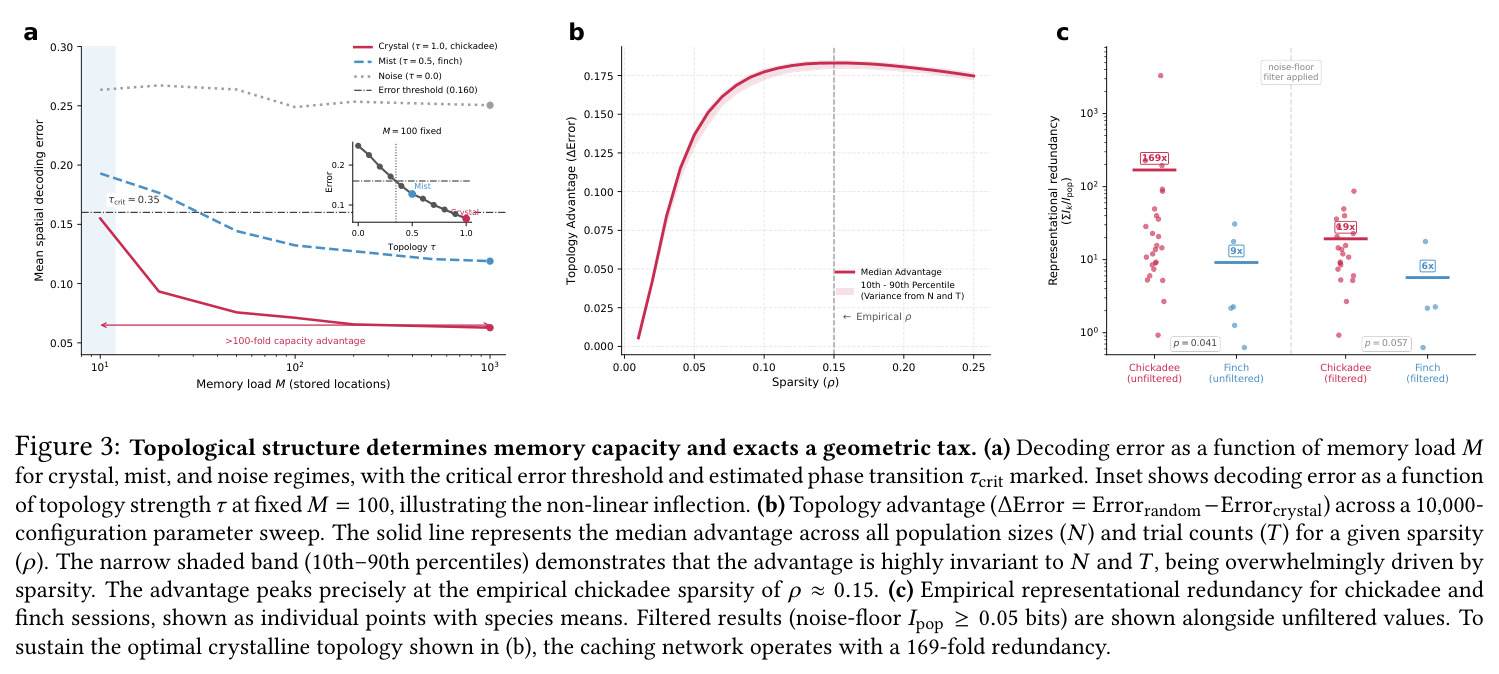
方法整体遵循'经验观察 → 指标量化 → 回路机制 → 计算验证'的闭环逻辑。先用 Shesha 几何稳定性指标在 Payne 2021 公开电生理数据集上对比山雀与草雀,再用主角度谱与本征维度量化兴奋-抑制群的正交互补,最后用 10000 组参数扫描的合成群体编码模型建立几何相变与记忆容量的因果链。直觉上,研究者把海马当作一个'固体物理系统',把记忆容量类比于相变温度之上的稳定晶格容量——晶体编码可承载 1000+ 个非干扰记忆,雾态编码在 10 个记忆处即崩溃,二者间存在 $\tau_{\text{crit}} \approx 0.35$ 的尖锐相变边界。
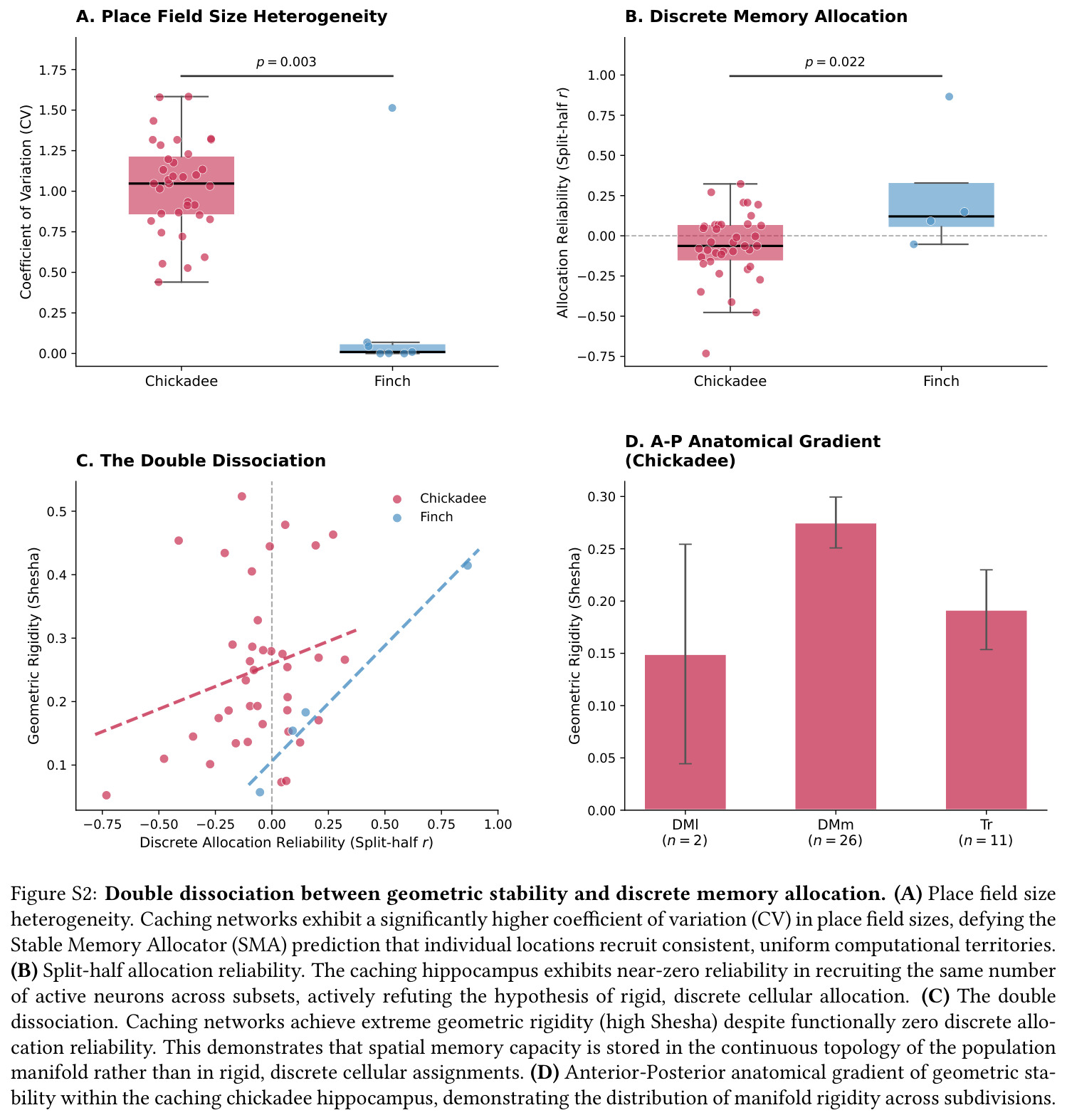
核心创新是把神经编码的'质量'从单神经元精度重定义为群体流形的拓扑刚性——具体由 Shesha 指标(分裂半 RDM 相关)与 Mantel 相关(神经-物理距离对应)共同刻画。与 Valiant 的离散分配模型相比,本文的'几何相变'框架主张高容量源于连续流形组织而非固定神经元分配,因此能解释为何山雀在分裂半分配可靠性近零($r=-0.064$)时仍维持极高几何稳定性($\text{Shesha}=0.245$)。
方法步骤详情
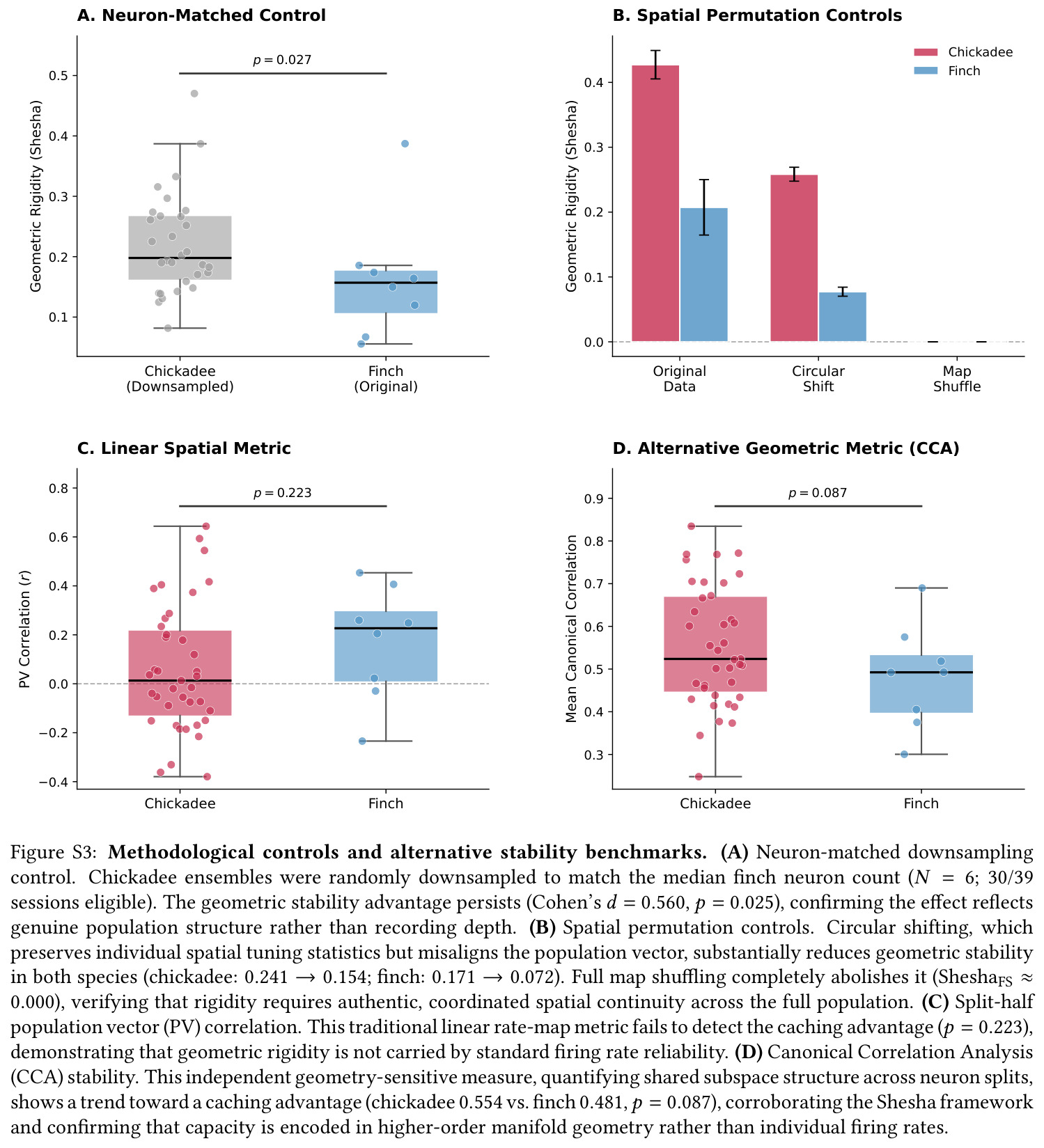
第一步,使用公开数据集(H. Payne 2021)中 39 个山雀 session 与 8 个草雀 session 的单单位放电,按开旷场位置构建每个 session 的群体向量并计算 RDM。第二步,对每个 session 实施 1000 次置换的 Mantel 检验,比较 RDM 距离与物理欧氏距离矩阵的 Spearman 相关,并以随机降采样至草雀中位数 $N=6$ 个神经元做匹配控制。第三步,计算 Shesha 几何稳定性——将神经元随机分半,分别计算两个 RDM 的相关,并验证圆形置换与图置换彻底消除稳定性以排除伪迹。第四步,按波形宽度区分兴奋(E)与抑制(I)细胞,分亚群计算 Shesha、$\text{bits/spike}$、Mantel $r$ 与时序互相关。第五步,对 $n=21$ 个 E-I 同时记录的 session 计算 E、I 群前三主成分子空间之间的三个主角度,并构造反相关对照计算参与比 $D$。第六步,构建合成 1D 圆形轨道吸引子网络($N=500, s=0.15$),扫描拓扑参数 $\tau \in [0,1]$(crystal/mist/noise 三档)与记忆负载 $M \in [10, 1000]$,解码误差为归一化圆周解码误差。第七步,对 $N, T, \rho$ 三维 10000 配置做参数扫描,计算 $\Delta_{\text{Error}} = E_{\text{random}} - E_{\text{crystal}}$ 并刻画其相对 $\rho$ 的非线性曲线。第八步,估计每 session 的表征冗余 $\text{redundancy} = \sum_i I_i / I_{\text{pop}}$,按 $I_{\text{pop}} \geq 0.05$ bits 的过滤阈值做主分析与稳健性分析。
技术新颖性
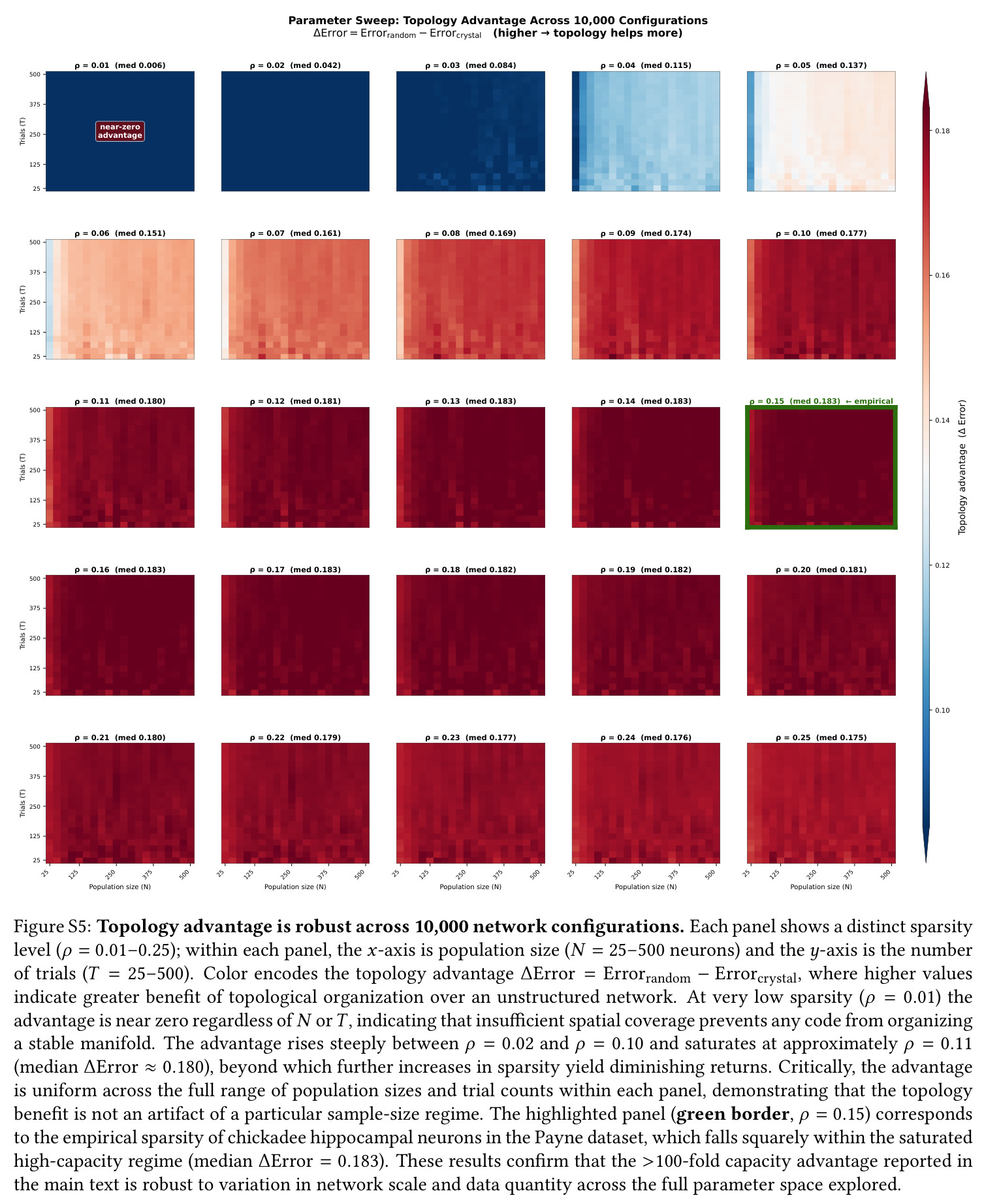
技术新颖性体现在三方面:(i) 把 Shesha 分裂半 RDM 稳定性指标引入到跨物种比较,并配合 Mantel 检验与下采样控制严格剥离神经元数效应;(ii) 把 E-I 回路机制量化为主角度谱($14°, 64°, 82°$)与本征维度扩展($\Delta D = +2.3$),从而把抽象的'抑制-去相关'概念落到可测量的子空间几何;(iii) 在 10000 配置扫描中证实拓扑优势 $\Delta_{\text{Error}}$ 主要由稀疏度 $\rho$(而非硬件规模 $N$ 或试验数 $T$)决定,且山雀经验稀疏度 $\rho \approx 0.15$ 正好落在饱和高优势区,这是以往单点模拟无法揭示的。
实验结果
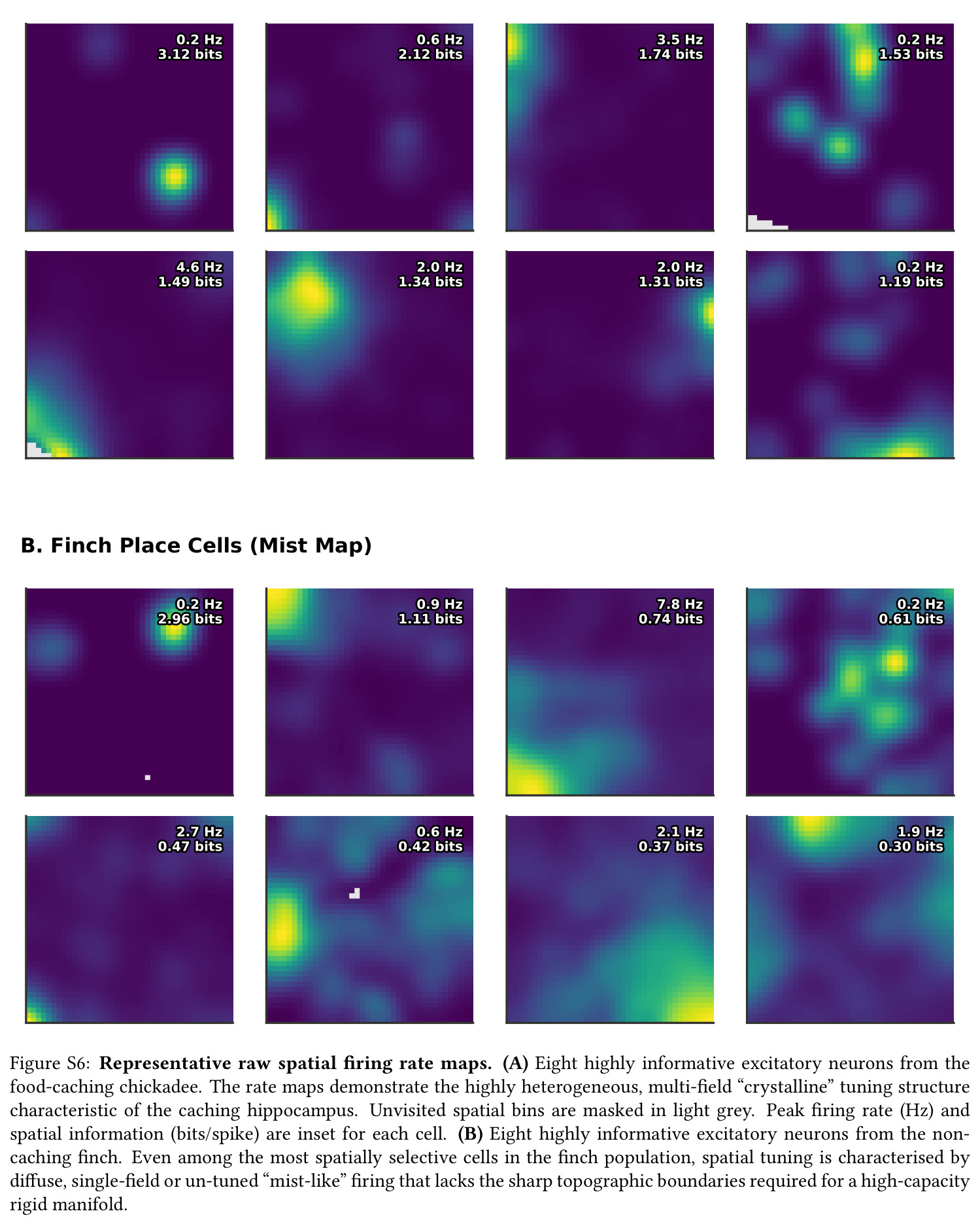
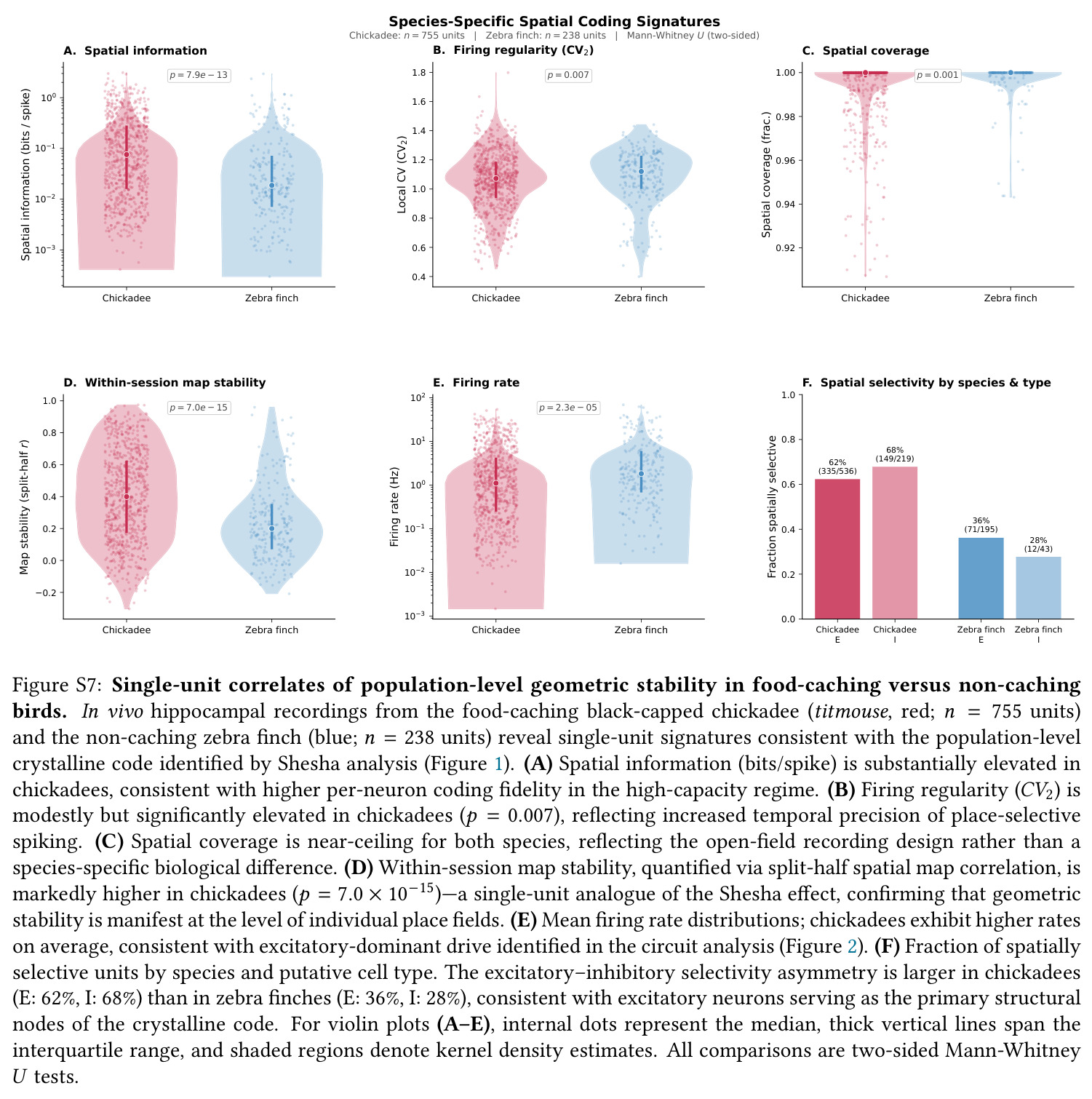
核心发现一:山雀海马维持显著更刚性的'晶体式'群体几何。Mantel $r$ 山雀均值为 $0.420$(39/39 session 显著),草雀仅为 $0.230$(8/8 显著),Mann-Whitney $p = 0.0012$;Shesha 几何稳定性山雀 $0.245$ vs 草雀 $0.166$,时间相干性山雀 $0.393$ vs 草雀 $0.209$,配对差异在 1000 次置换下全部显著。核心发现二:分裂半分配可靠性呈反向双分离——山雀均值 $r = -0.064$(接近零),显著低于草雀 $0.264$($p = 0.022$ 单尾),且山雀位置野尺寸变异系数高达 $1.03$ vs 草雀 $0.23$($p = 0.003$),证伪离散神经元分配假说。核心发现三:兴奋-抑制回路呈'一共享两正交'架构,前三主角度均值为 $14°, 64°, 82°$(后者接近随机独立子空间期望值 $\sim 83°$),后验概率 $P(E>I) > 97.5\%$。核心发现四:抑制群使本征维度从 $D = 7.5$ 扩展至 $D = 9.8$(+2.3 维),反相关对照不产生扩展,证伪简单减法抑制机制。核心发现五:在合成网络中晶体编码在 $M = 1000$ 仍维持亚阈值误差,雾态编码在 $M = 10$ 即崩溃,容量优势 >100 倍;10000 配置扫描显示 $\Delta_{\text{Error}}$ 主要由稀疏度 $\rho$ 决定,在 $\rho \geq 0.11$ 时饱和于 $\approx 0.183$,山雀经验 $\rho \approx 0.15$ 正落在此饱和高优势区。核心发现六:山雀经验表征冗余中位数为 12 倍(过滤后),非过滤均值 19.3 倍,与 169 倍理论预测同向但偏低(源于细胞外采集的天花板效应)。
查看结构化数据
| 任务 | 指标 | 本文 | 基线 | 提升 |
|---|---|---|---|---|
| 跨物种 Mantel 物理-神经距离对应 | Spearman $r_{\text{Mantel}}$(中位数,39 vs 8 sessions) | chickadee $r = 0.420$ | finch $r = 0.230$ | Mann-Whitney $p = 0.0012$,效应量为中等偏大且 100% session 显著 |
| 群体几何稳定性(Shesha 分裂半 RDM 相关) | Shesha FS 分数 | chickadee $0.245$ | finch $0.166$ | 相对提升约 47.6%,且在随机降采样至 $N=6$ 后仍保留 Cohen's $d = 0.560$ |
| 时序相干性(within-session PV cross-correlation) | 相关系数 | chickadee $0.399$ | finch $0.241$ | 约 1.66 倍提升,$p < 0.001$,代表晶体编码'同一张印刷地图反复读取' |
| 分裂半位置细胞分配可靠性(Valiant SMA 指标) | 分裂半 Pearson $r$ | chickadee $r = -0.064$ | finch $r = 0.264$ | 反向双分离:山雀几何更优但分配可靠性显著更低,$p = 0.022$ 单尾 |
| 合成网络晶体 vs 雾态容量阈值 | 可维持亚阈值解码误差的 $M$ 上界 | crystal 编码可至 $M = 1000$ | mist 编码在 $M = 10$ 即超阈值 | 容量提升 >100 倍,临界拓扑阈值 $\tau_{\text{crit}} \approx 0.35$ |
| 兴奋-抑制本征维度扩展 | 参与比 $D$ | E+I 联合 $D = 9.8$ | E 单独 $D = 7.5$;反相关对照 $D = 7.5$ | 净增 +2.3 维,95% 贝叶斯 CI $[0.024, 0.143]$ 排除零效应 |
局限与改进
作者明确承认的局限包括:(i) E-I 回路分析仅基于 $n=13$ 个 session,贝叶斯可信区间虽排除零但仍属欠样本,频率派推断需要更大同步记录数据集;(ii) 计算模型因高维'测度集中'问题只能在 1D 圆形轨道上演示,无法直接扩展至 2D 开旷场;(iii) 经验对比仅依赖单一公开数据集、两物种,缺乏跨物种或多脑区的趋同验证;(iv) 经验冗余中位数 12 倍远低于理论 169 倍,作者解释为细胞外采样上限效应但未提供独立验证。此外作者未明确讨论的潜在问题包括:分裂半 RDM 相关本身可能对小样本($N \approx 6-21$)神经元数高度敏感,圆周解码误差作为唯一容量指标是否对非空间任务有泛化性,$\tau_{\text{crit}} \approx 0.35$ 是否对其他物种具有普适性,以及'几何税'是否在能量代谢层面也有代价——这些都是值得延伸讨论的方向。
独立分析的弱点
独立审视本文仍有若干可改进之处。其一,Shesha 与 Mantel 都属于二阶统计量,对非线性几何敏感度有限,建议加入拓扑数据分析(TDA 的持久同调)以更严格刻画'晶体 vs 雾态'的拓扑差异。其二,E-I 主角度谱虽漂亮但只在 $n=21$ session 上观察,且缺乏光遗传或药理学操纵抑制群后的因果验证——若能在山雀上做抑制群急性沉默并观察 Shesha 是否崩溃,将是直接因果证据。其三,10000 配置扫描虽证实拓扑优势由稀疏度主导,但 $N$ 与 $T$ 的边际贡献可能在更大的生物合理范围内($N=10^4$-$10^5$)才显现,目前 $N \leq 500$ 的扫描不足以完全排除'堆神经元也行'的论点。其四,论文未控制两种鸟的探索策略差异——若山雀在觅食中采用更序列化的访问模式(与 Raju 2024 序列假说相符),其 Mantel 优势可能部分归因于行为而非神经机制。其五,圆周解码误差阈值的 $0.640$ 是事后定义,缺乏独立心理物理学锚定。
未来方向
作者提出的未来方向包括:在更大同步 E-I 数据集上做频率派复制、把 2D 拓扑扩展到可处理的网格或低维流形模型、从其他储食物种(如鸦科)和哺乳动物海马收集趋同证据、以及检验内嗅皮层网格尺度差异是否构成上游因果基础。基于本文成果可进一步延伸的研究包括:(i) 把'几何相变'概念迁移到人工神经网络中作为归纳偏置,例如在 Hopfield/Transformer 注意力机制中引入拓扑刚性正则;(ii) 探索发育可塑性——幼年山雀是否通过练习强化几何相变;(iii) 把 $\tau_{\text{crit}} \approx 0.35$ 作为阿尔茨海默病早期诊断的'几何崩溃'标志物;(iv) 在强化学习框架下把'几何税'形式化为表征效率与抗噪性的 Pareto 前沿;(v) 用闭环虚拟现实范式在线操纵山雀的 E-I 平衡并测 Shesha 因果响应。
复现评估
论文在方法学上具备较高复现性:数据来源为 Payne 等 2021 公开数据集,所有自定义代码已在 GitHub(github.com/prashantcraju/hippocampal-stability)开源,详细方法见附录 A 并引用所有非参数置换次数(10000)、Mantel 置换次数(1000)、Bayesian 估计的可信区间计算方式。复现所需算力较低——电生理分析可在单核 CPU 完成,10000 配置的合成网络扫描是主要瓶颈,在单 GPU 上数小时可完成;但 1D 圆形轨道网络已经逼近当前解析极限,2D 扩展需要全新的数学工具。复现难度中等偏高,主要挑战不在算力而在概念正确性:研究者需精确实现 Shesha 分裂半指标、主角度谱的 SVD 分解、以及参与比的特征值计算,任何一处偏差都会显著影响山雀 vs 草雀的小效应量比较。
论文图表