通用多模态蛋白质设计实现化学的DNA编码 General Multimodal Protein Design Enables DNA-Encoding of Chemistry
DISCO多模态扩散模型实现蛋白质序列-结构协同设计,无需预定义活性位点即可设计出新酶
前置知识
蛋白质语言模型
蛋白质语言模型是预训练在大量蛋白质序列上的深度神经网络,通过自监督学习如掩码语言建模捕捉蛋白质序列的统计规律和氨基酸间的依赖关系。它将氨基酸序列映射到连续嵌入空间,能够提取进化信息和结构-功能关系,为下游任务提供丰富的特征表示。在蛋白质设计中,pLM可以编码部分序列信息,帮助模型理解合理的序列模式。
DISCO使用冻结的蛋白质语言模型来编码部分序列,这是跨模态回收机制的核心组件,理解pLM对于理解DISCO如何整合序列先验知识至关重要。
扩散模型
扩散模型是一种生成式深度学习框架,通过逐步添加噪声将数据转换为简单分布如高斯分布,然后学习反向去噪过程来从噪声中生成新样本。对于连续数据如3D坐标,使用标准去噪分数匹配,对于离散数据如氨基酸序列,使用掩码扩散语言模型。DISCO的关键创新是将这两种扩散过程统一到一个联合生成框架中。
DISCO的核心就是多模态扩散,理解扩散模型的基本原理对于掌握DISCO的工作机制至关重要。
协同设计
协同设计是指同时优化多个相互依赖的设计变量,在蛋白质设计中特指同时设计氨基酸序列和三维结构,而不是先生成结构再设计序列或反之。由于序列决定结构、结构影响功能,两者紧密耦合,协同设计能够在生成早期就利用两种模态的信息,产生更优的设计方案。DISCO的跨模态回收机制实现了真正的协同设计。
协同设计是DISCO与现有方法的本质区别,也是其能够发现新活性位点几何结构的关键。理解这一概念有助于把握论文的核心贡献。
催化机理
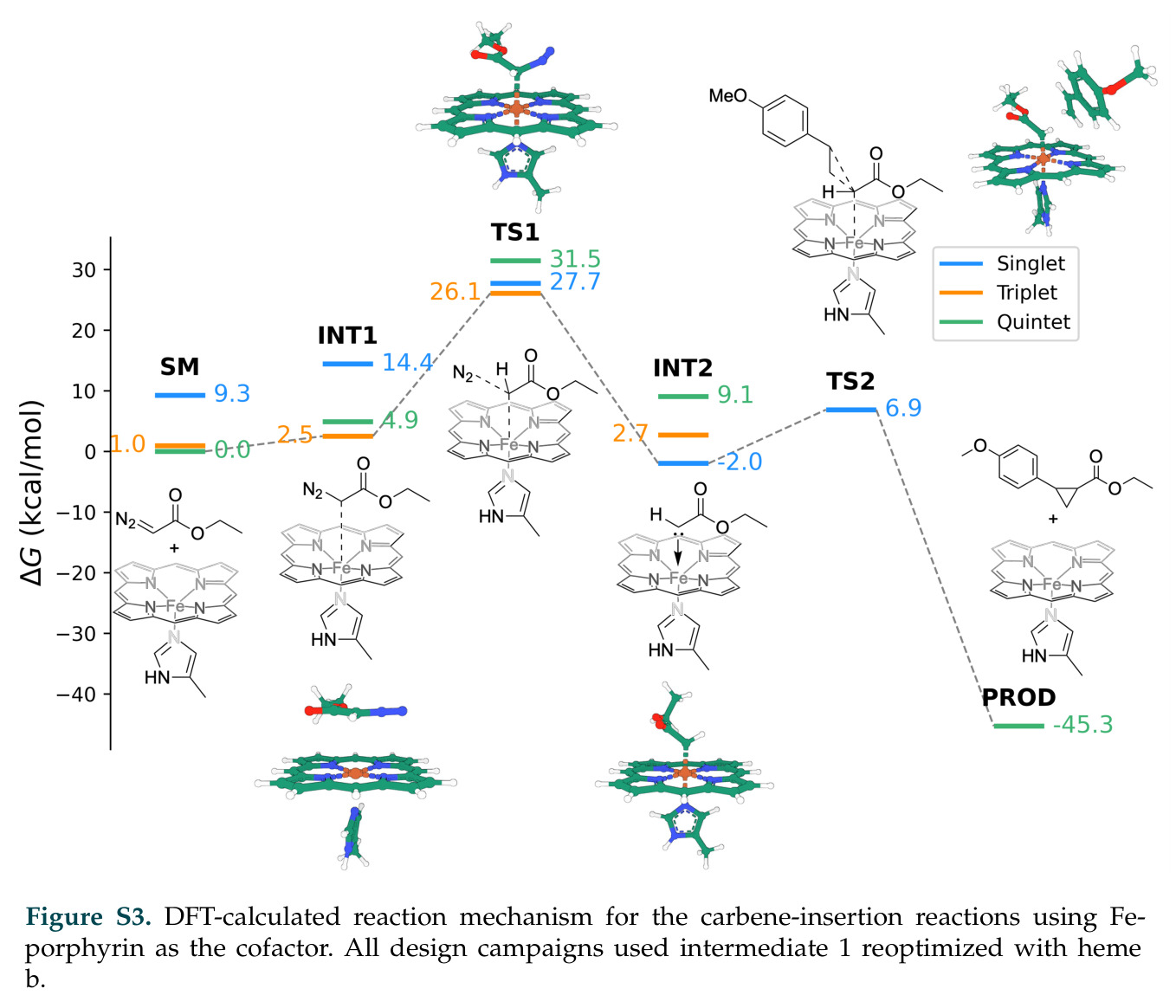
催化机理描述酶如何降低反应能垒,包括活性位点的几何排列、催化残基的电子相互作用、过渡态的稳定等。对于卡宾转移反应,关键中间体是铁-卡宾物种,其形成通常是决速步,能垒超过20千卡每摩尔。DISCO的创新之处在于不需要预先指定催化残基或精确的过渡态几何,只需基于反应中间体条件就能设计出功能酶。
理解催化机理有助于明白为什么DISCO的方法比传统的基于theozyme的方法更通用、更灵活。
研究动机
现有蛋白质设计方法面临两个根本限制:第一,它们严重依赖预定义的、固定的活性位点残基或theozyme几何排列,这排除了那些没有已知 motif 或精确机制的反应,阻碍了新活性位点几何结构的发现;第二,现有的计算流程大多是序贯的:先生成骨架再设计匹配的序列,由于序列和结构共同决定功能,这种解耦的范式无法在关键的结构形成阶段利用基于序列的目标,反之亦然。对于自然界中不存在的新颖化学反应,找到合适的酶作为定向进化的起点仍然极其困难,通常需要基于化学直觉筛选有限的已知骨架,这个过程不仅耗时,而且受限于进化已经探索过的空间。
本文的目标是本文的目标是开发一个通用的多模态框架,能够同时从头设计蛋白质序列和三维结构,无需预定义残基 motif,并且可以基于任意生物分子进行条件生成。具体来说,DISCO旨在仅基于反应中间体而非完整的过渡态模型就能设计出具有多样化、可进化活性位点的血红素酶,催化自然界不存在的卡宾转移反应,这些酶的活性应达到或超过定向进化典型起点的水平,甚至可以与广泛进化的变体相媲美。
与已有工作不同的是,本文的独特切入点在于完全摒弃了预定义催化残基或theozyme的传统做法,而是让模型的协同折叠机制采样与正在构建的蛋白质相容的反应中间体构象。这意味着DISCO可以在不被人类关于催化所需残基或固定过渡态几何的假设所约束的情况下,探索催化解决方案。此外,DISCO的协同生成范式允许序列和结构相互感知和调整,这是序贯方法无法实现的。这种基于中间体而非过渡态的条件策略对于机制不确定性使得theozyme构建变得不可能的新颖化学反应尤其有价值。
核心方法
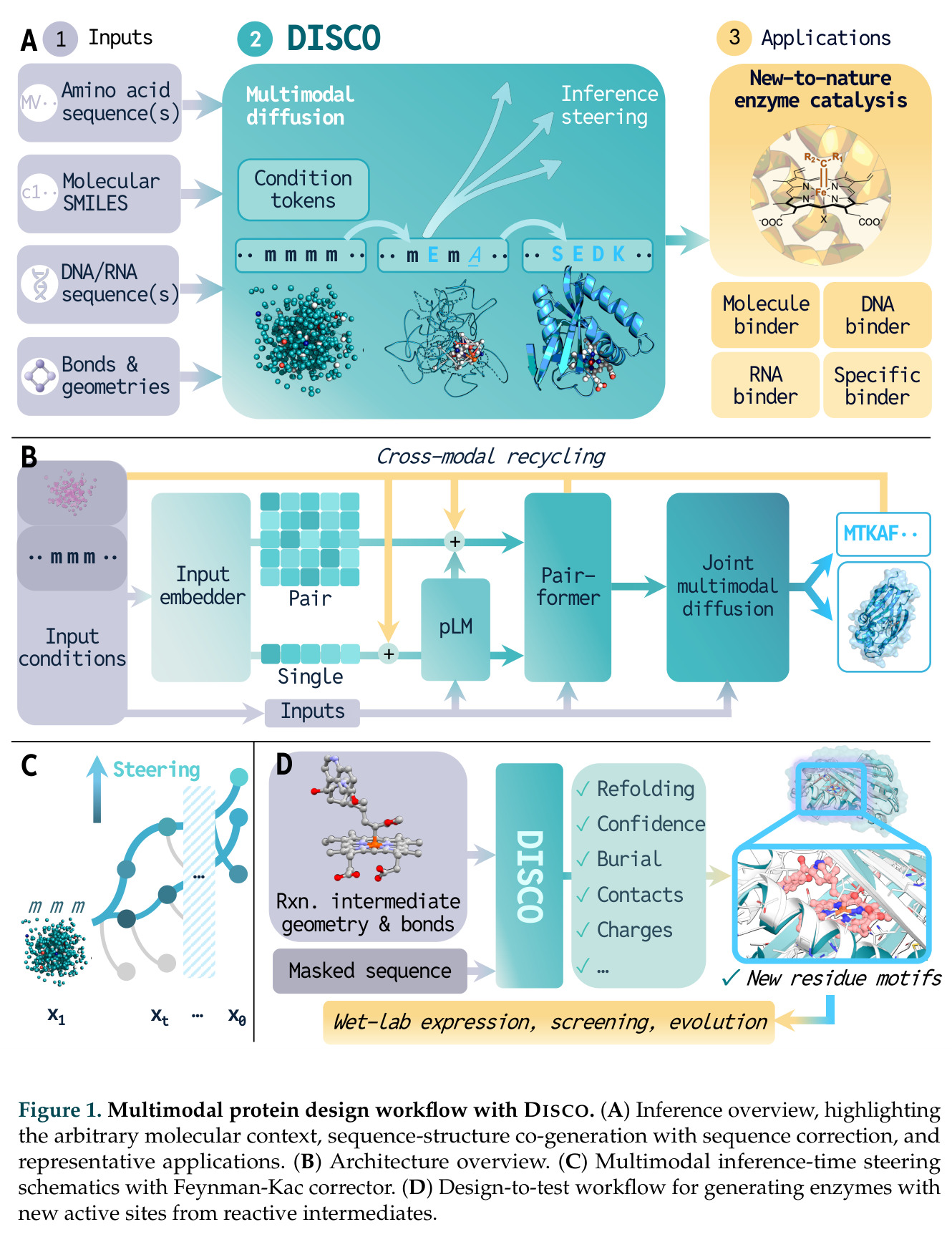
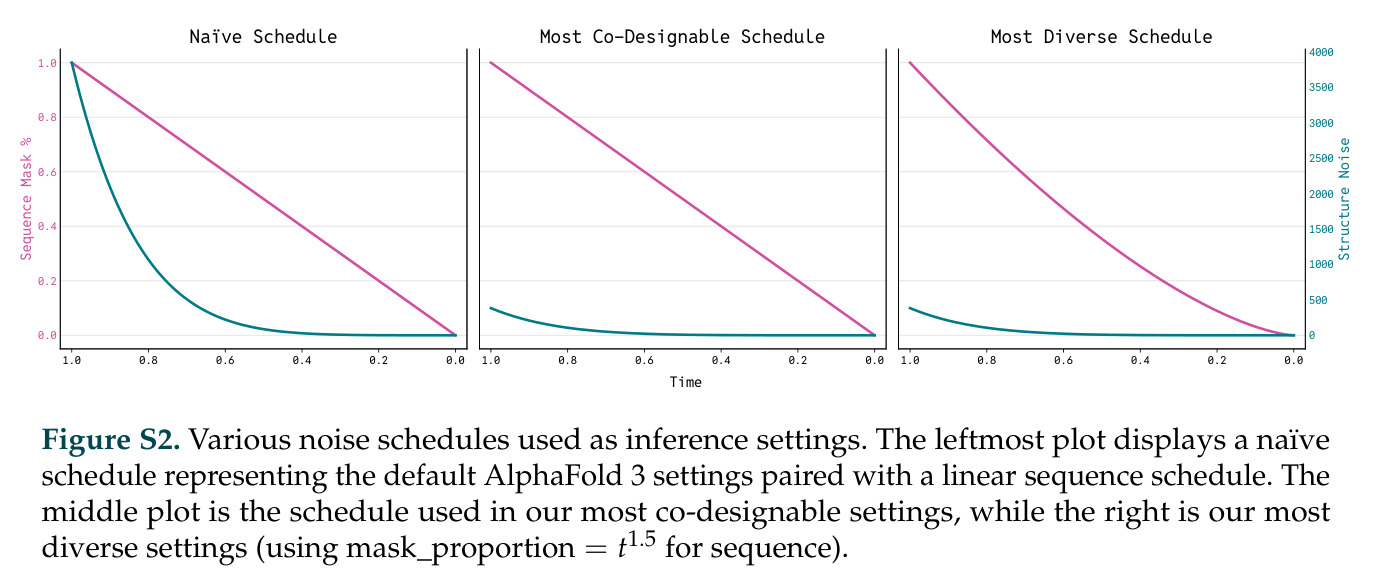
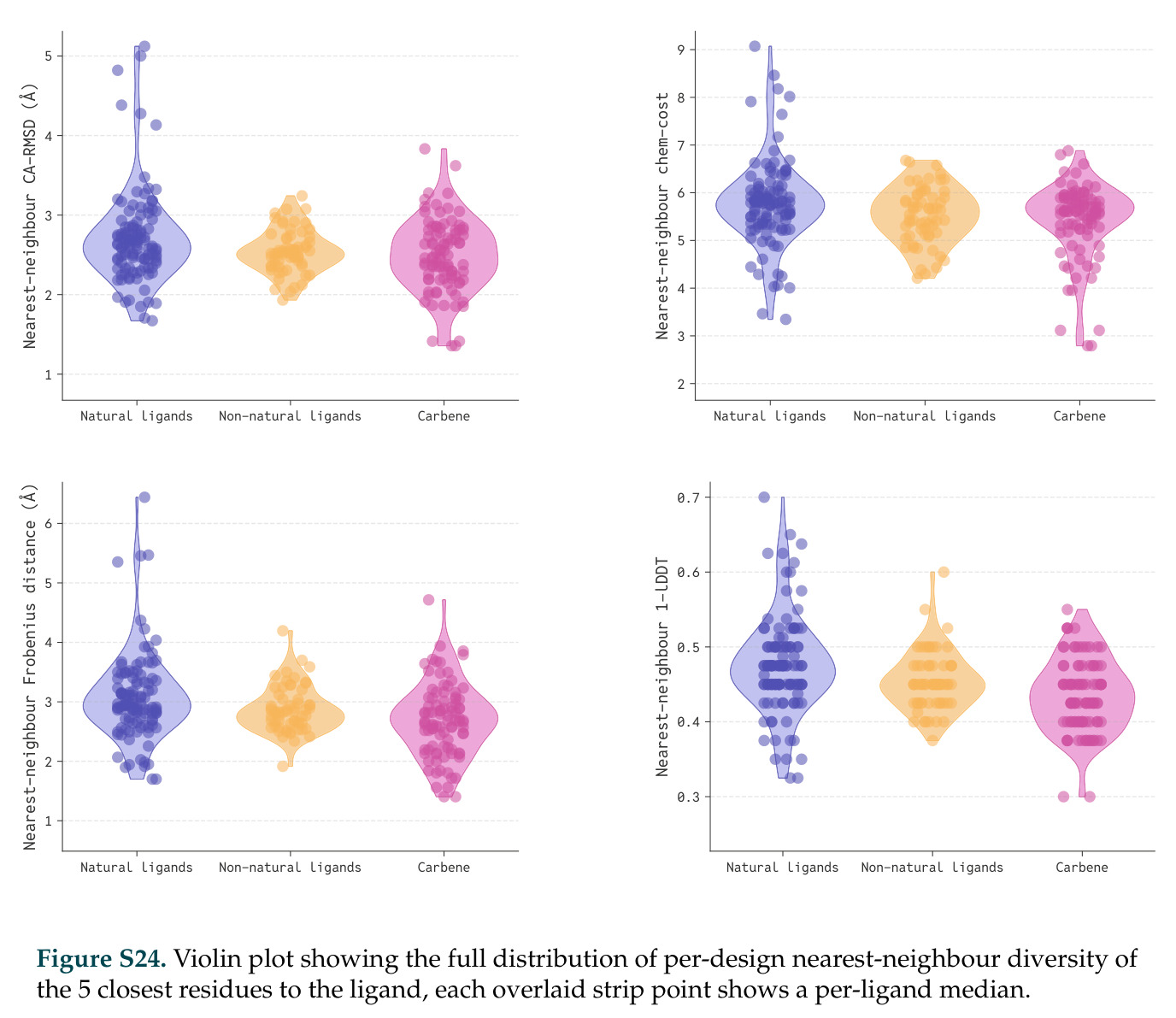
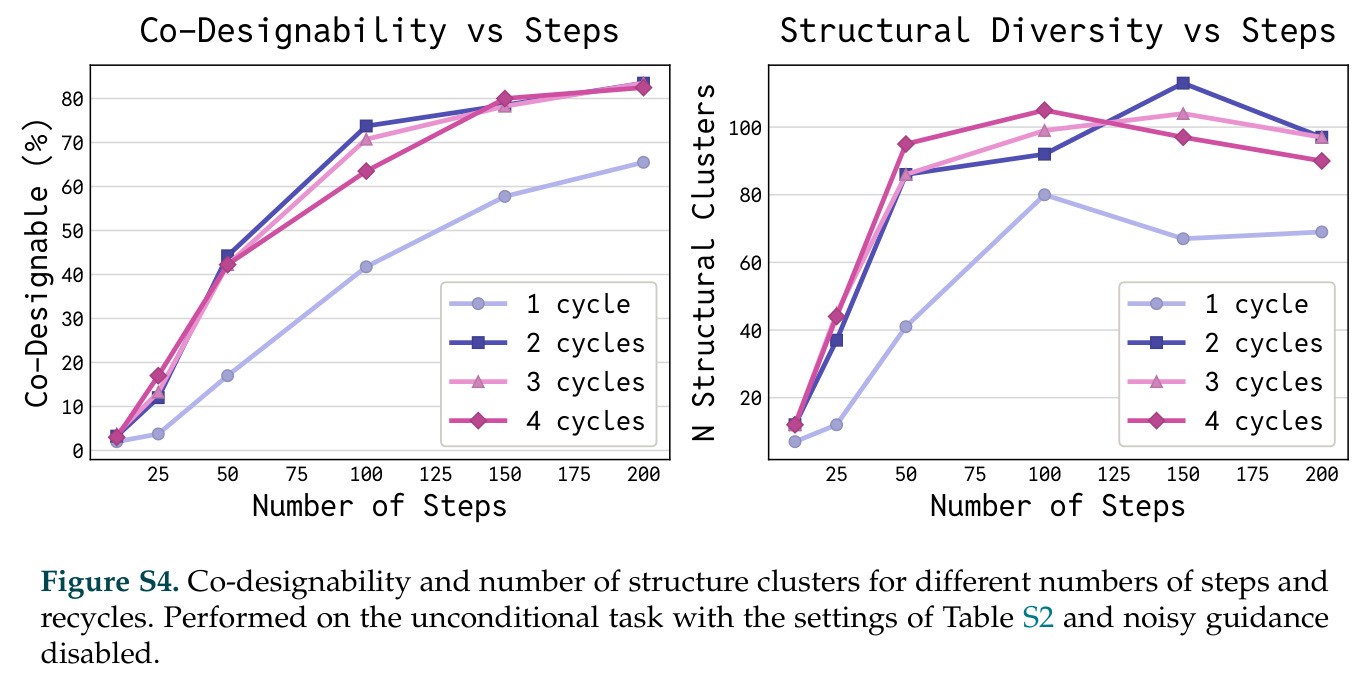
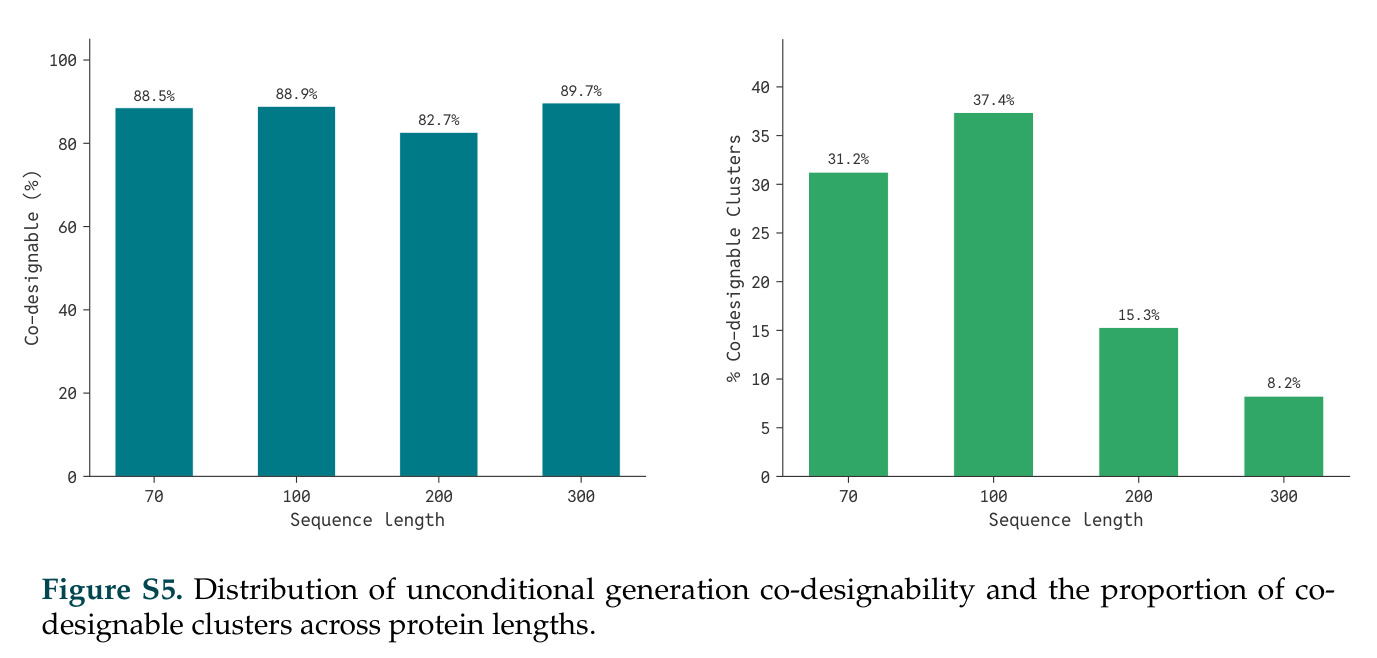
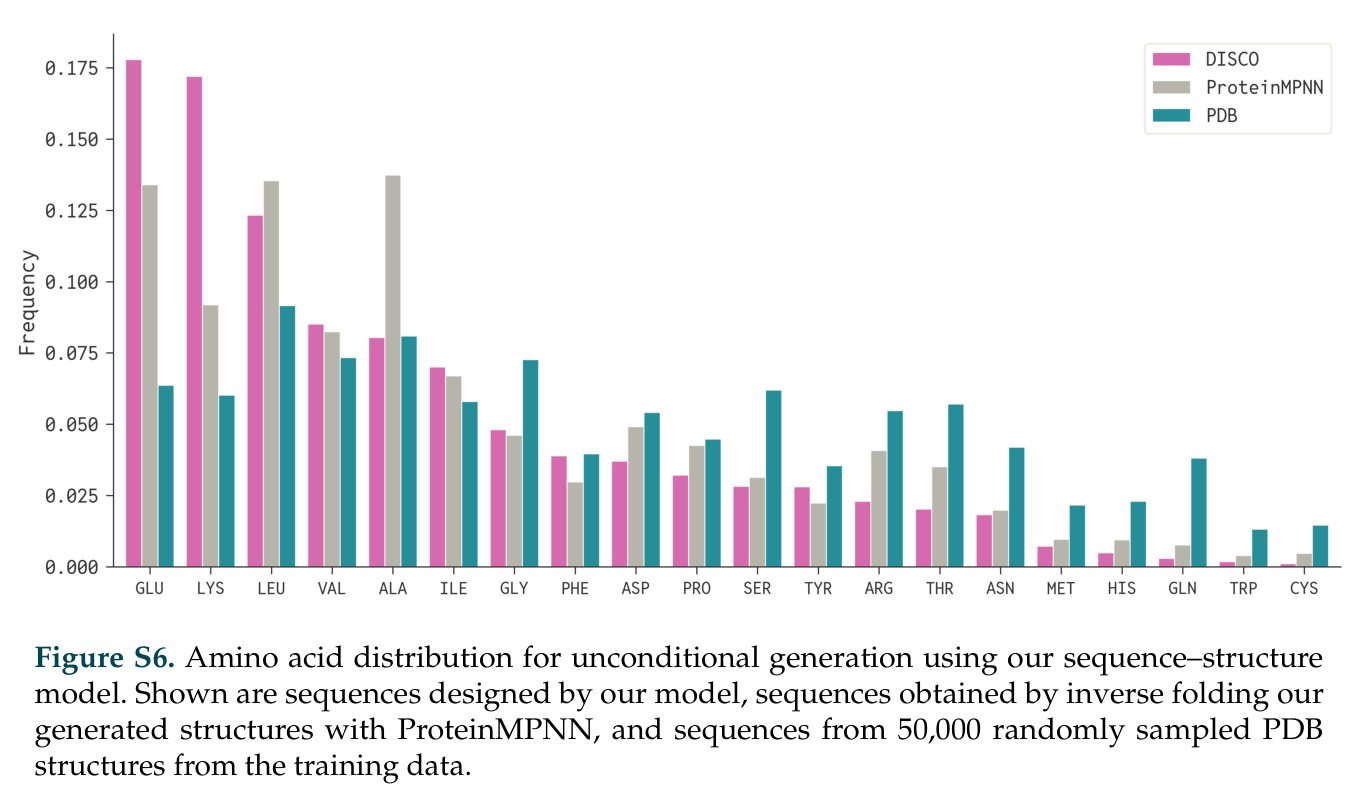
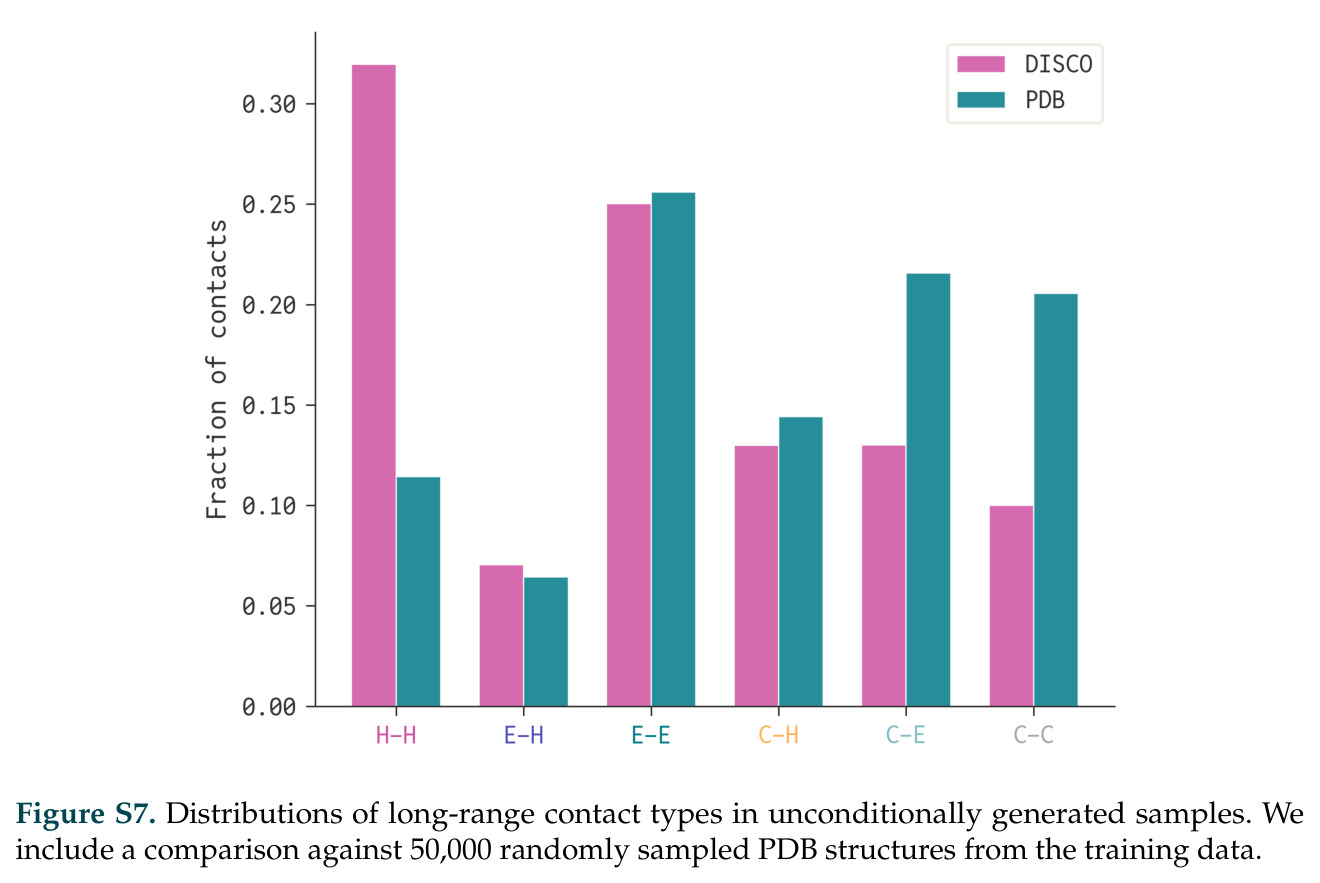
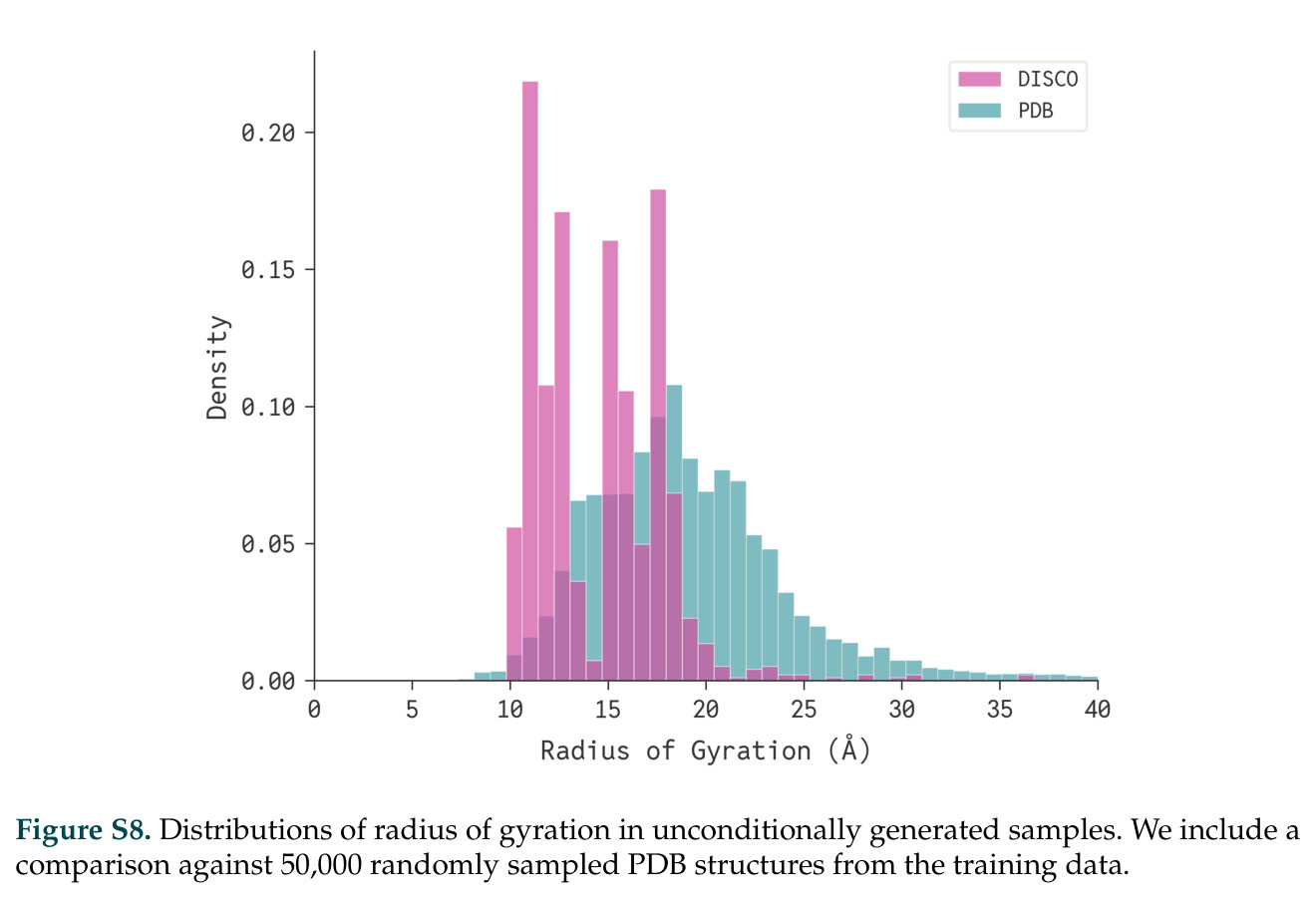
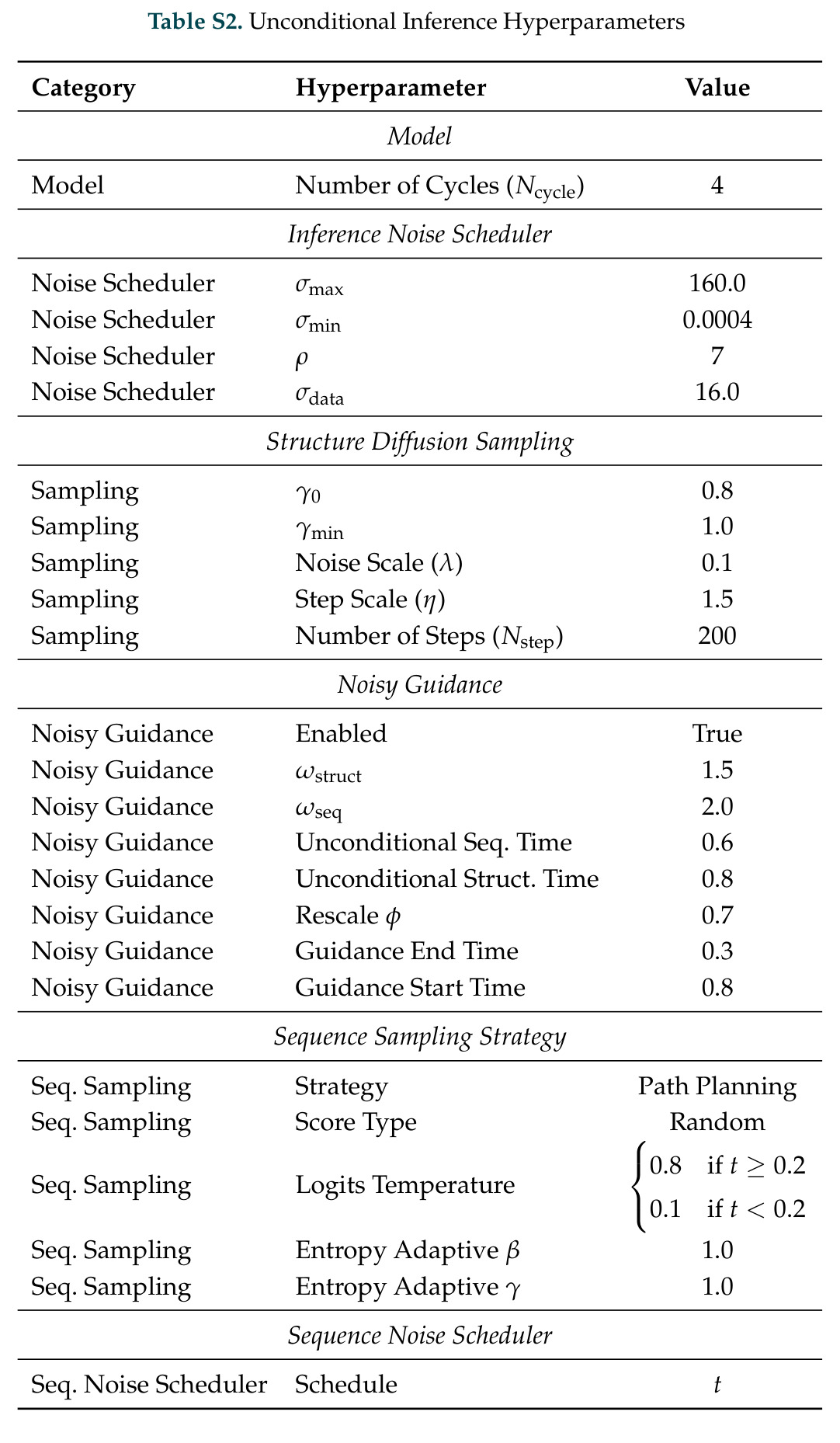
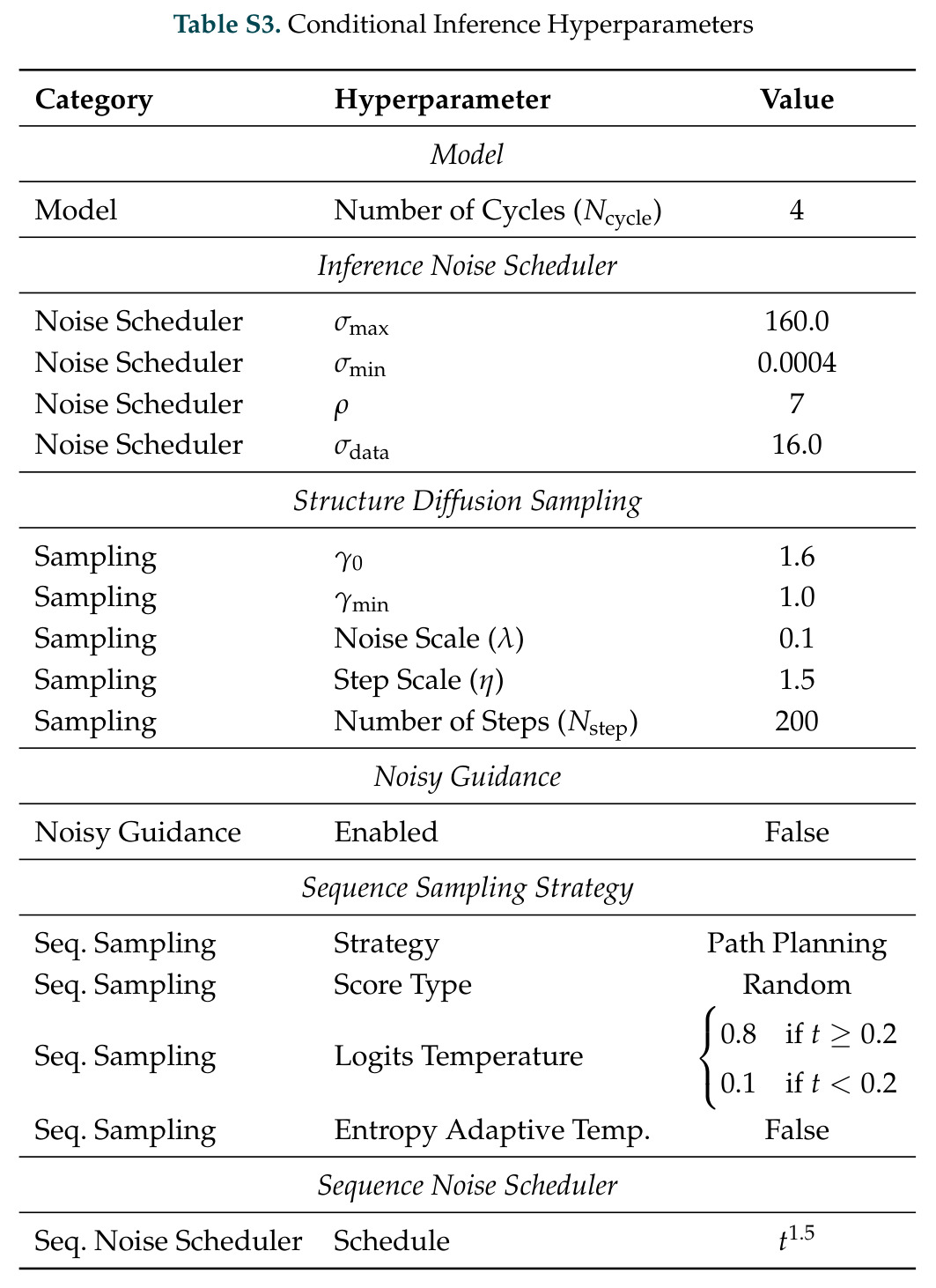
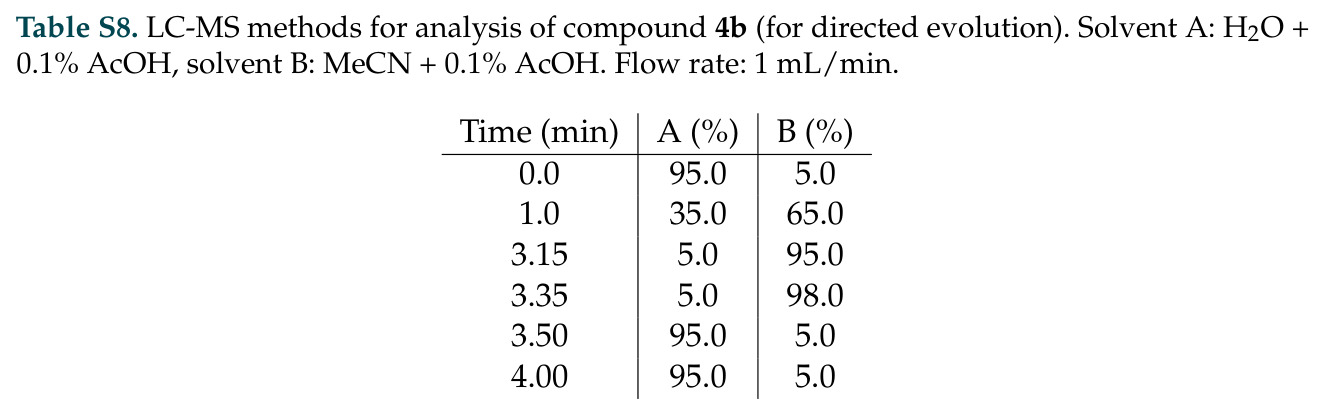
DISCO的整体思路是将离散的氨基酸序列和连续的3D原子坐标建模为联合分布,通过统一的生成过程进行去噪。模型训练一个单一的深度神经网络,该网络使用掩码离散扩散过程处理序列,同时利用连续扩散处理3D原子坐标。关键创新是跨模态回收机制,在每个生成步骤中,模型同时基于当前预测的干净序列和结构、以及当前加噪的序列和结构这四种编码进行条件生成,确保序列预测 informed by 正在出现的结构特征,结构预测也适应正在演化的序列身份。推理策略方面,引入了序列自校正机制和序列温度机制来显著提高协同设计能力,在无条件单体设计中,约90%的生成序列可以重折叠到其设计骨架结构的2埃RMSD范围内。
DISCO与已有方法的最本质区别在于:真正的多模态协同生成,同时生成序列和结构,通过跨模态回收实现双向信息流,而非序贯的先生成后匹配;任意生物分子条件生成,将非蛋白质分子以与蛋白质相同的方式表示,并让它们与蛋白质在整个生成轨迹中协同折叠,模型会调整条件生物分子的坐标以捕获分子相互作用诱导的构象变化;无需预定义 motif,仅基于反应中间体就能设计功能酶,而不是基于预先指定的催化残基几何;推理时扩展,通过Feynman-Kac校正器框架,可以同时用序列和结构信号进行推理时缩放,直接在生成过程中整合目标属性,而不是依赖低效的生成-过滤方法。
方法步骤详情
DISCO方法的完整步骤分为训练和推理两个阶段。训练阶段:从Protein Data Bank收集蛋白质序列-结构对数据,无需特殊过滤避免偏差;对序列应用掩码扩散过程,随机掩码部分氨基酸,对3D坐标应用连续扩散,逐步添加高斯噪声;通过独立的单模态扩散损失训练网络,坐标使用标准去噪分数匹配损失,序列使用掩码扩散语言模型损失;跨模态回收机制在每个步骤注入四种编码:当前预测的干净序列和结构,以及当前加噪序列和结构。推理阶段:输入目标分子如反应中间体的SMILES和几何;从噪声中逐步去噪,同时解开序列掩码;在每步应用自校正机制,用结构预测修正序列预测;应用序列温度机制,平滑过度自信token的分布;可选地使用Feynman-Kac校正器进行推理时引导,如FKC-MM用于多模态奖励倾斜增加二硫键含量或阳离子-派相互作用,FKC-SG用于特异性引导结合目标分子同时避免结构相似的decoy;最终输出协同设计的序列-结构对,用ESMFold或Chai-1验证重折叠质量。
技术新颖性
DISCO的技术新颖性体现在多个方面:首先,它首次实现了真正的多模态蛋白质扩散,将离散序列扩散和连续结构扩散统一到一个框架中,并通过跨模态回收机制实现真正的协同设计;其次,它提出了任意生物分子条件生成的通用方法,将非蛋白质分子作为第一类公民处理,并在生成过程中进行协同折叠,这突破了现有方法只处理固定、预定义原子支架的限制;第三,它引入了Feynman-Kac校正器框架在多模态设置中的两种新应用:FKC-MM允许用定义在离散序列和连续结构上的联合奖励函数进行奖励倾斜,FKC-SG通过采样倾斜分布鼓励样本在目标模型下可能同时惩罚在off-target模型下可能,从而实现特异性结合设计;第四,实验验证方面,它是首个仅基于反应中间体而非预定义活性位点或theozyme就成功设计出功能酶的工作,这些酶催化自然界不存在的卡宾转移反应,活性超过工程化酶的典型起点。
实验结果
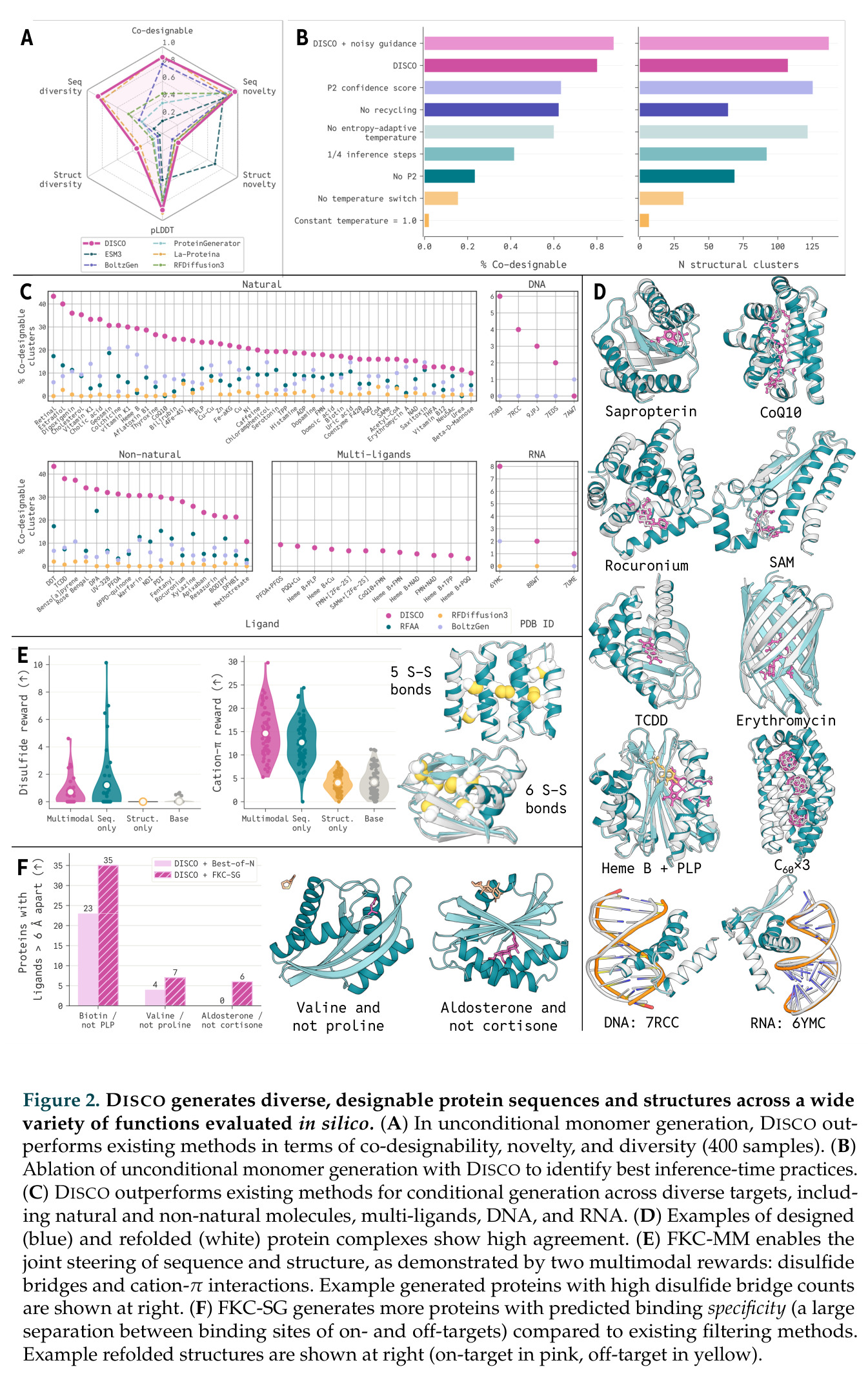
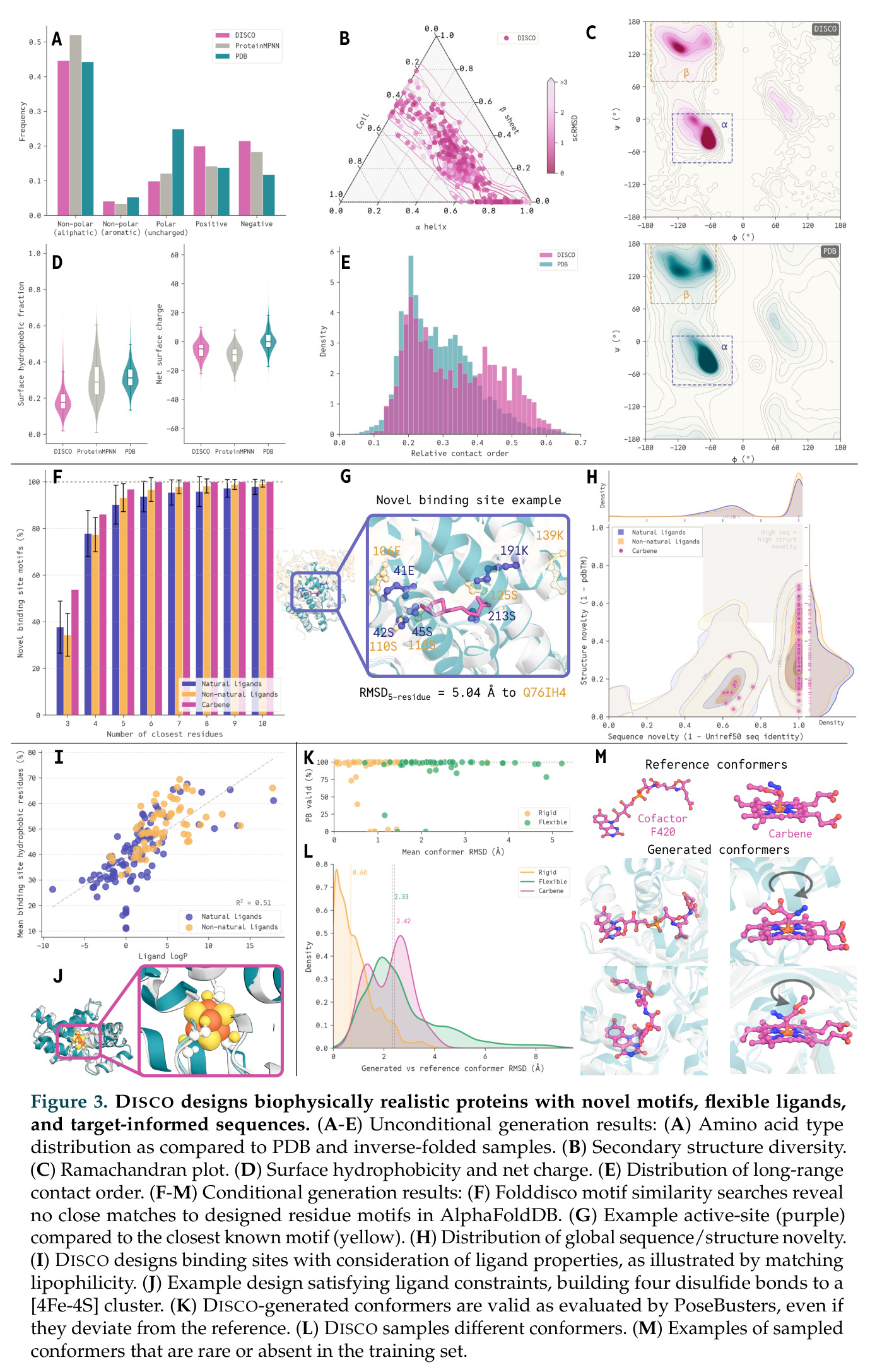
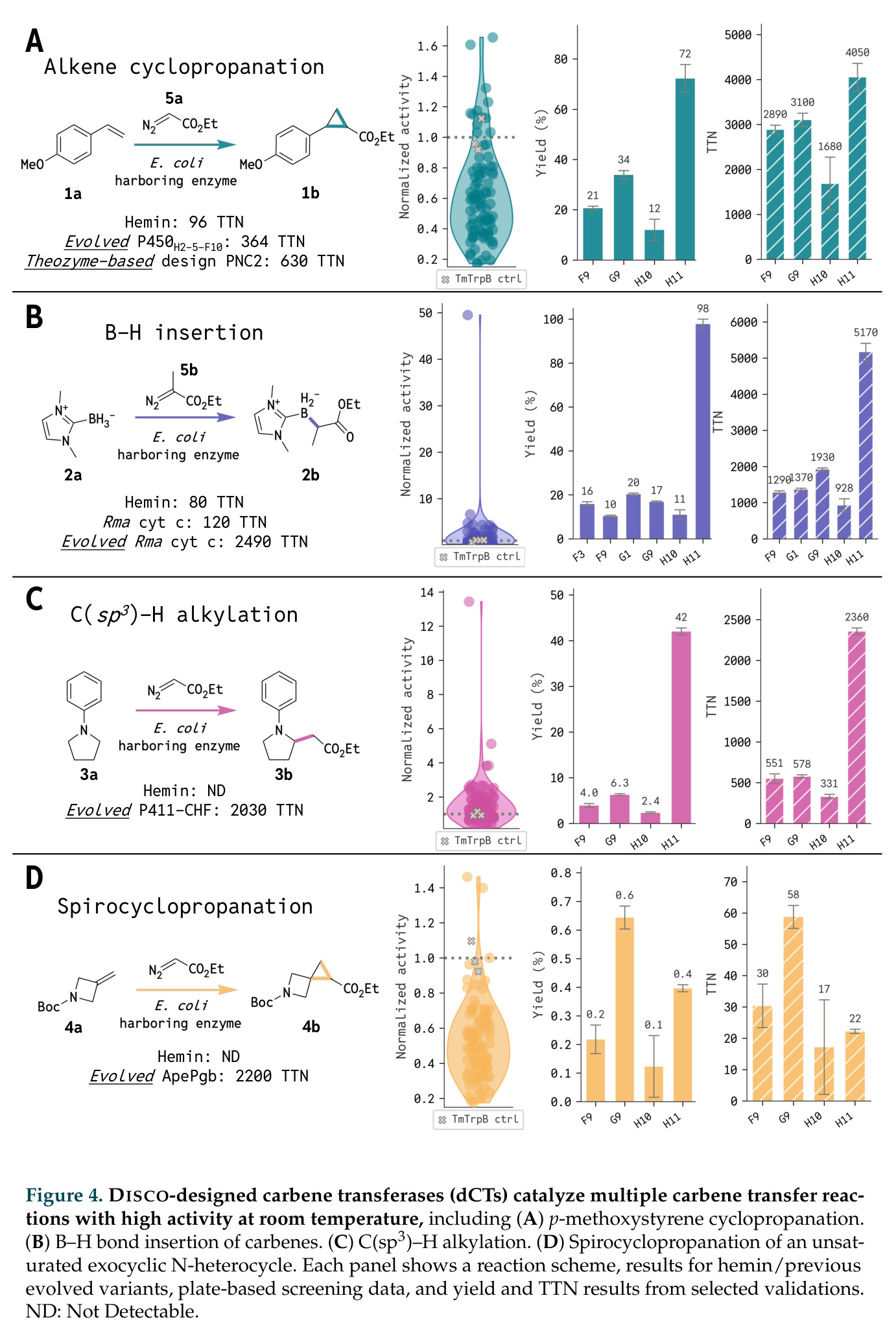
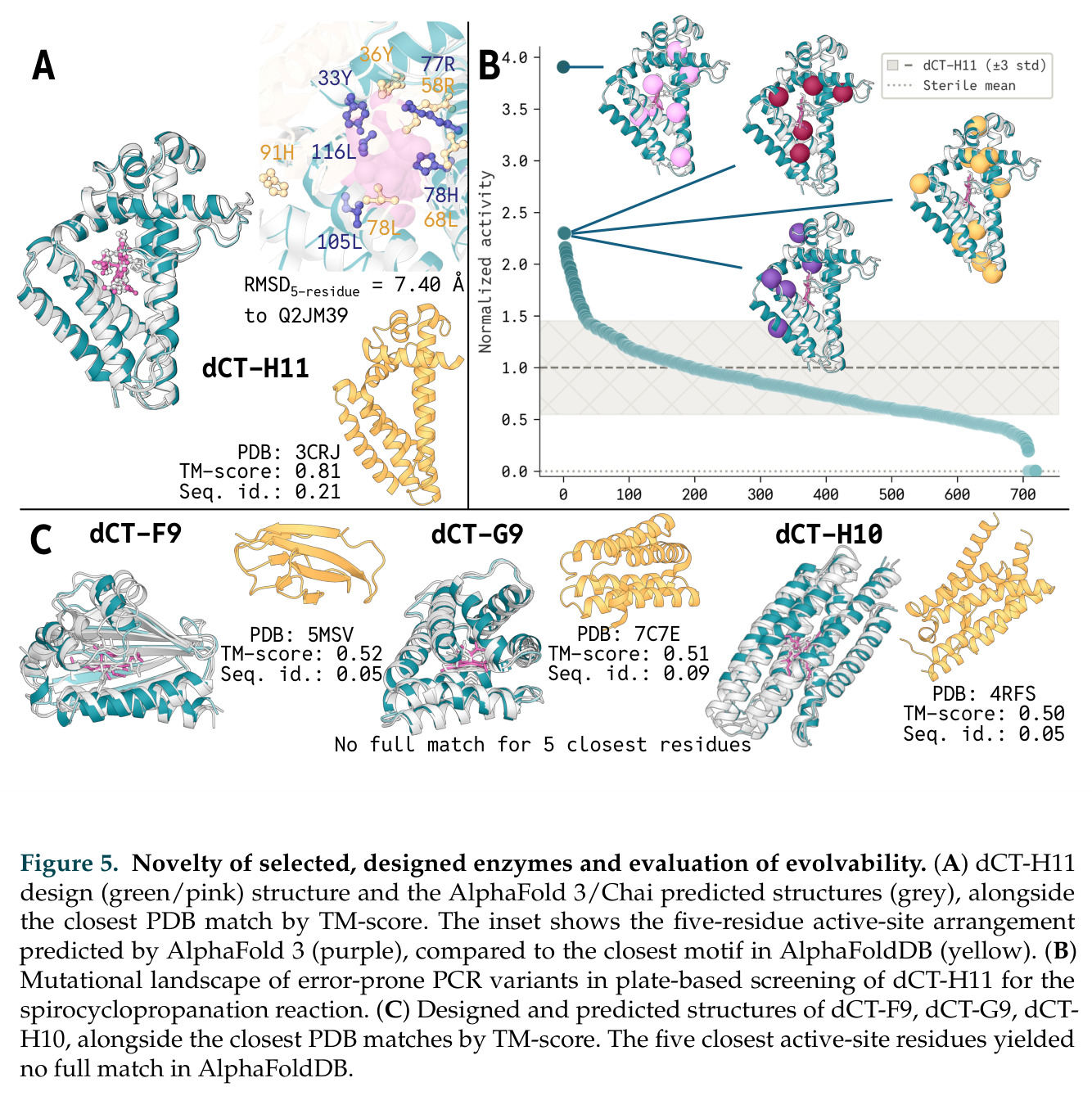
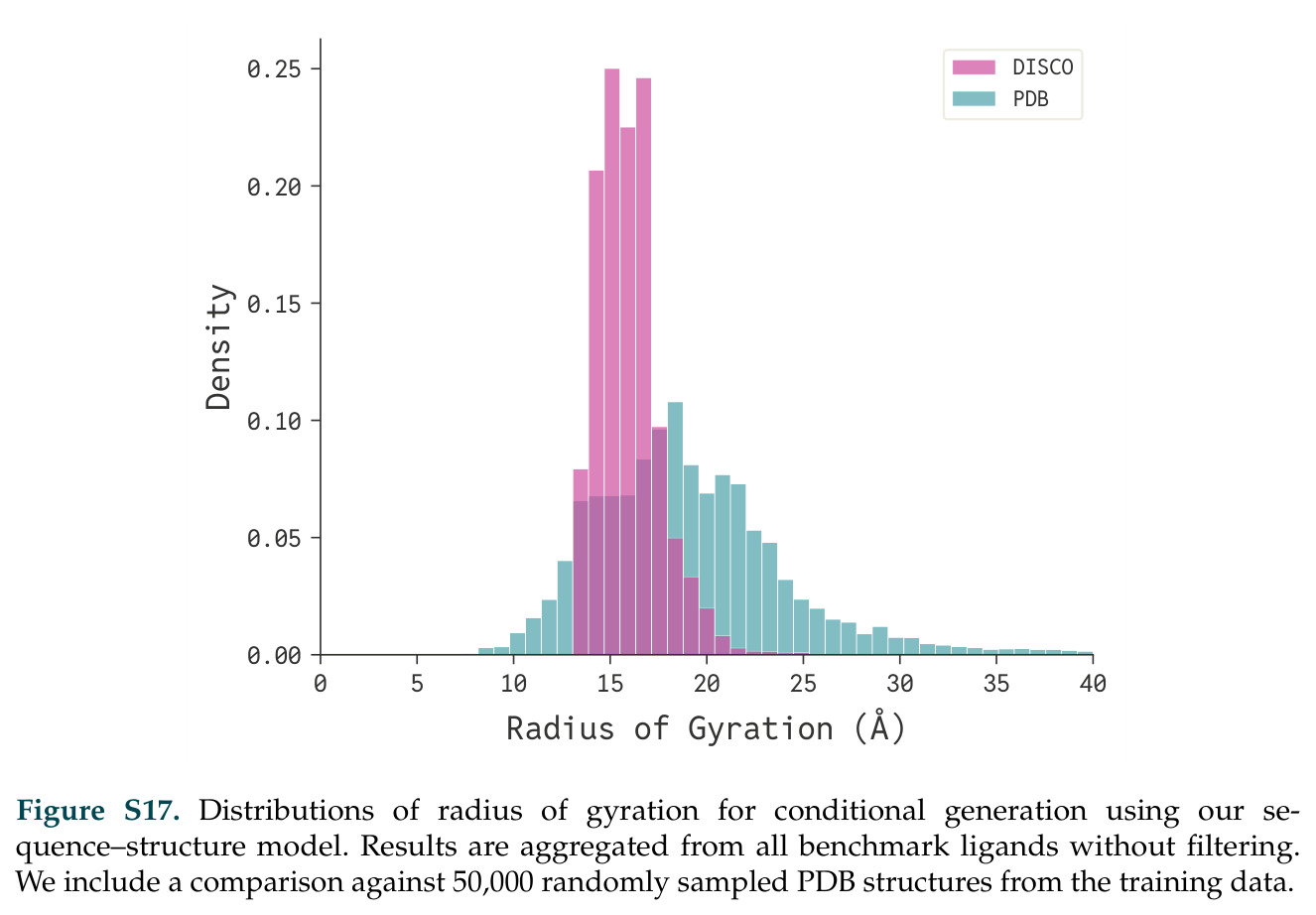
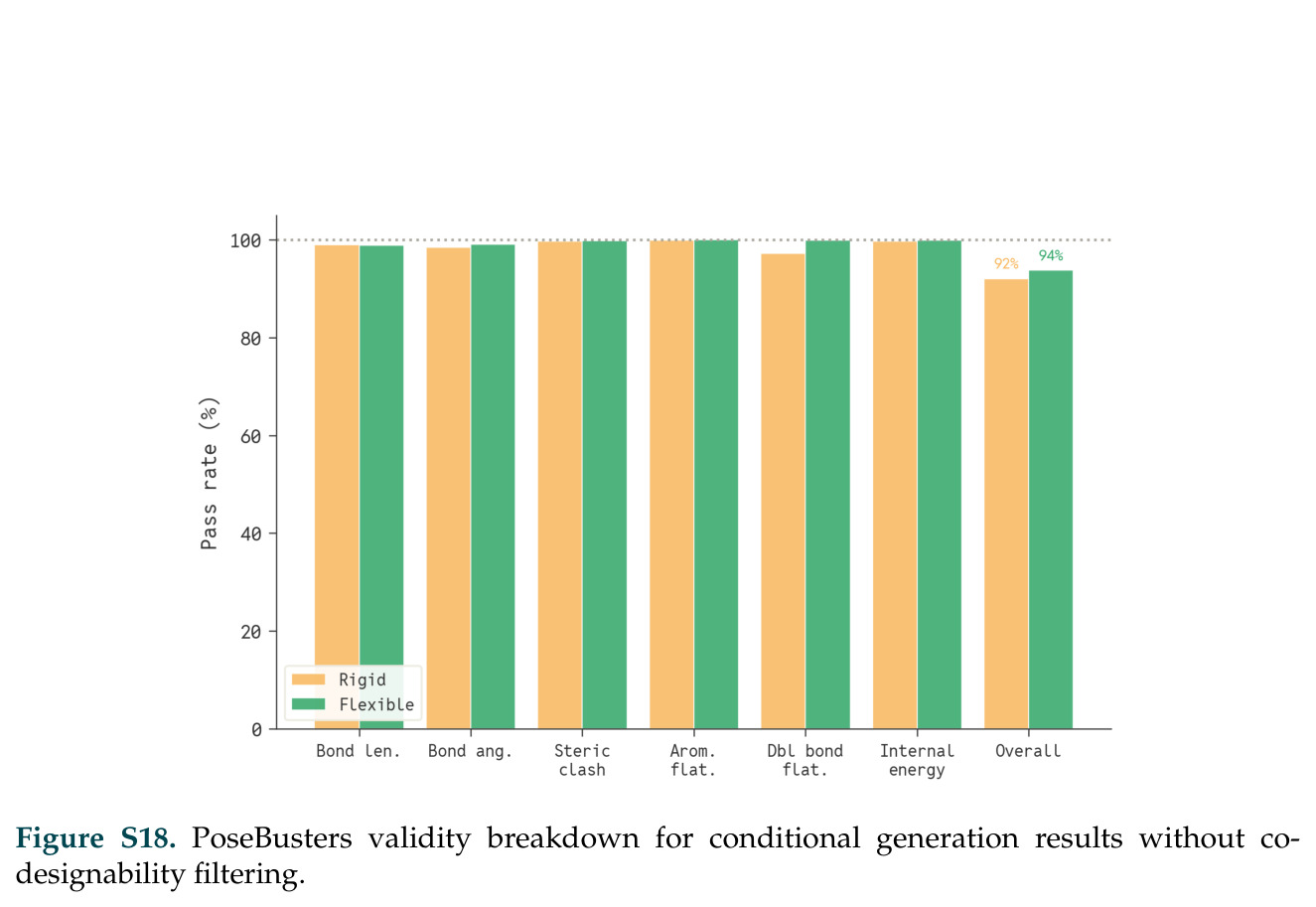
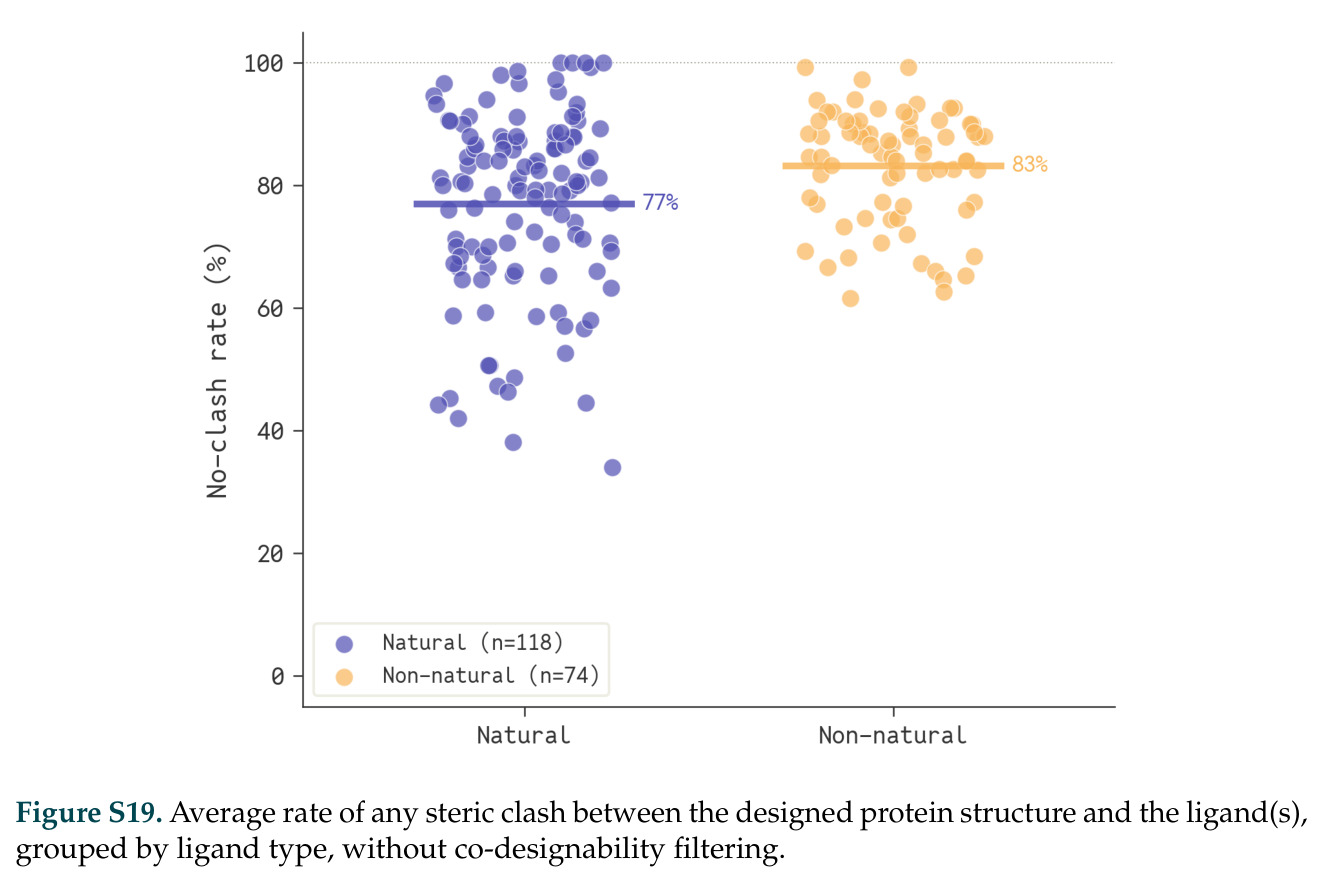
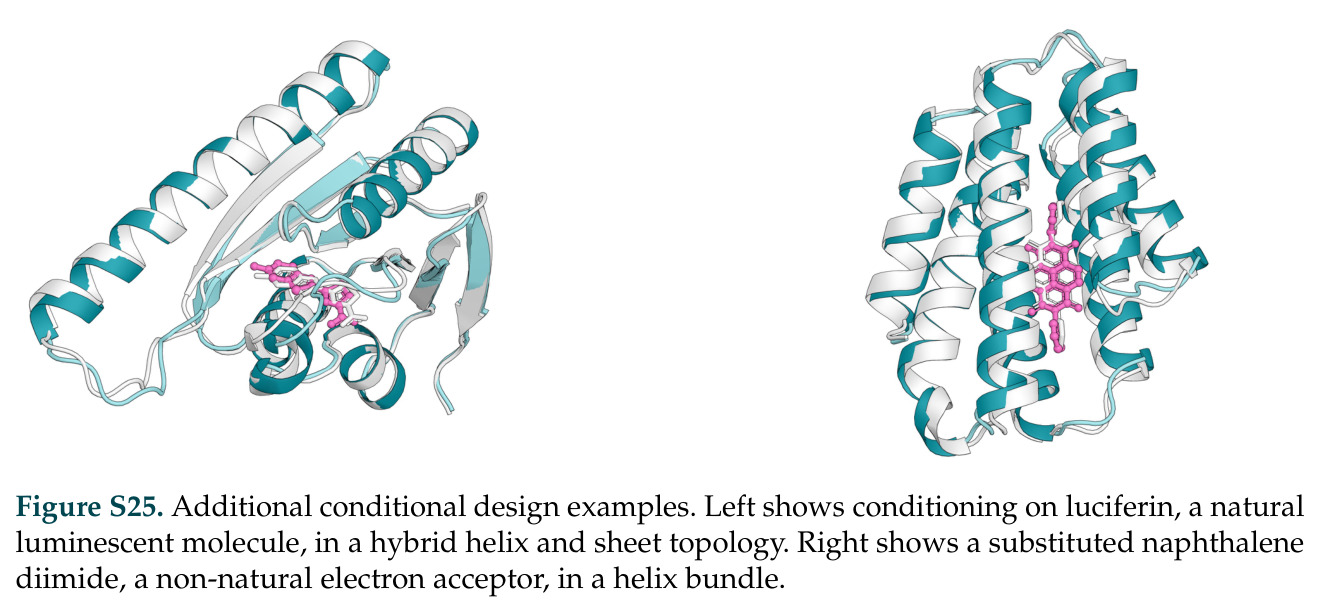
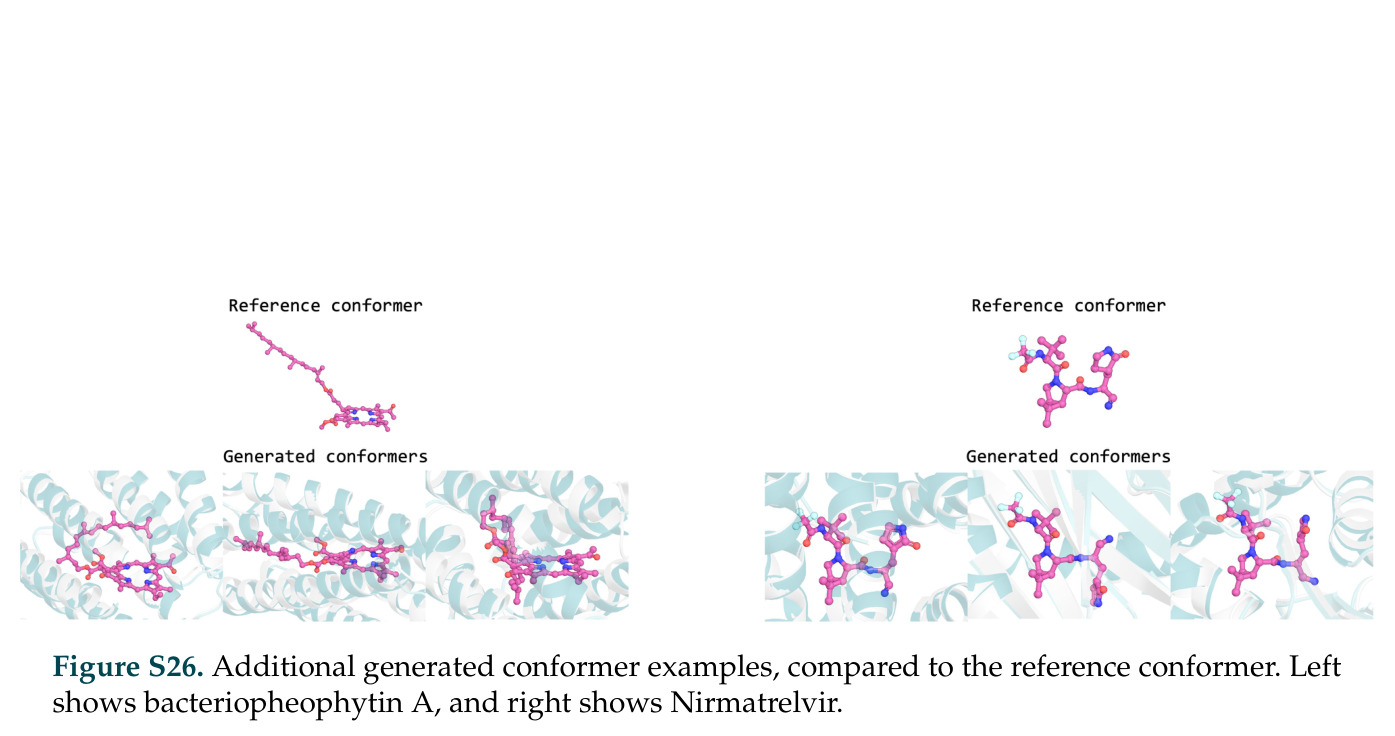
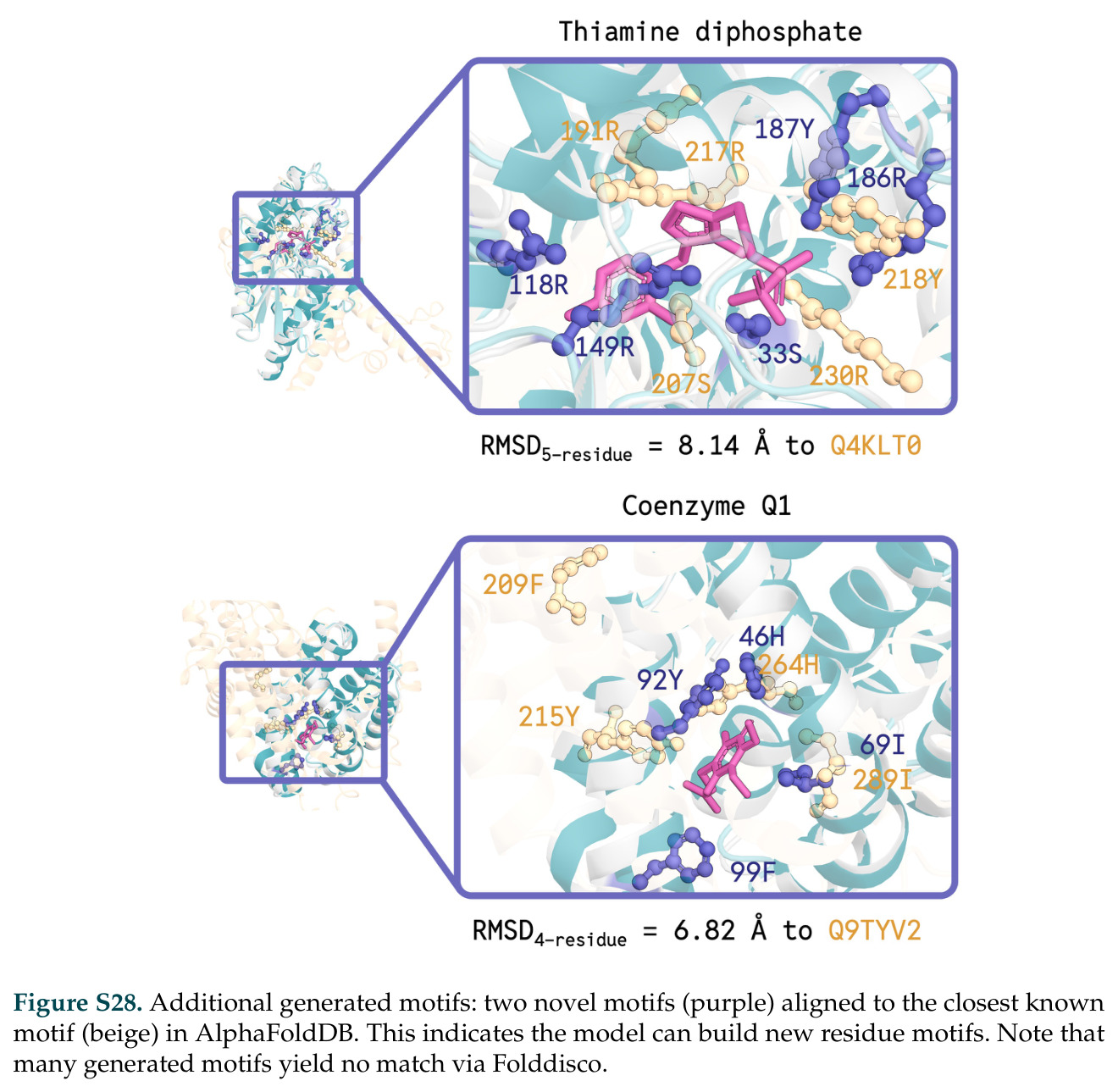
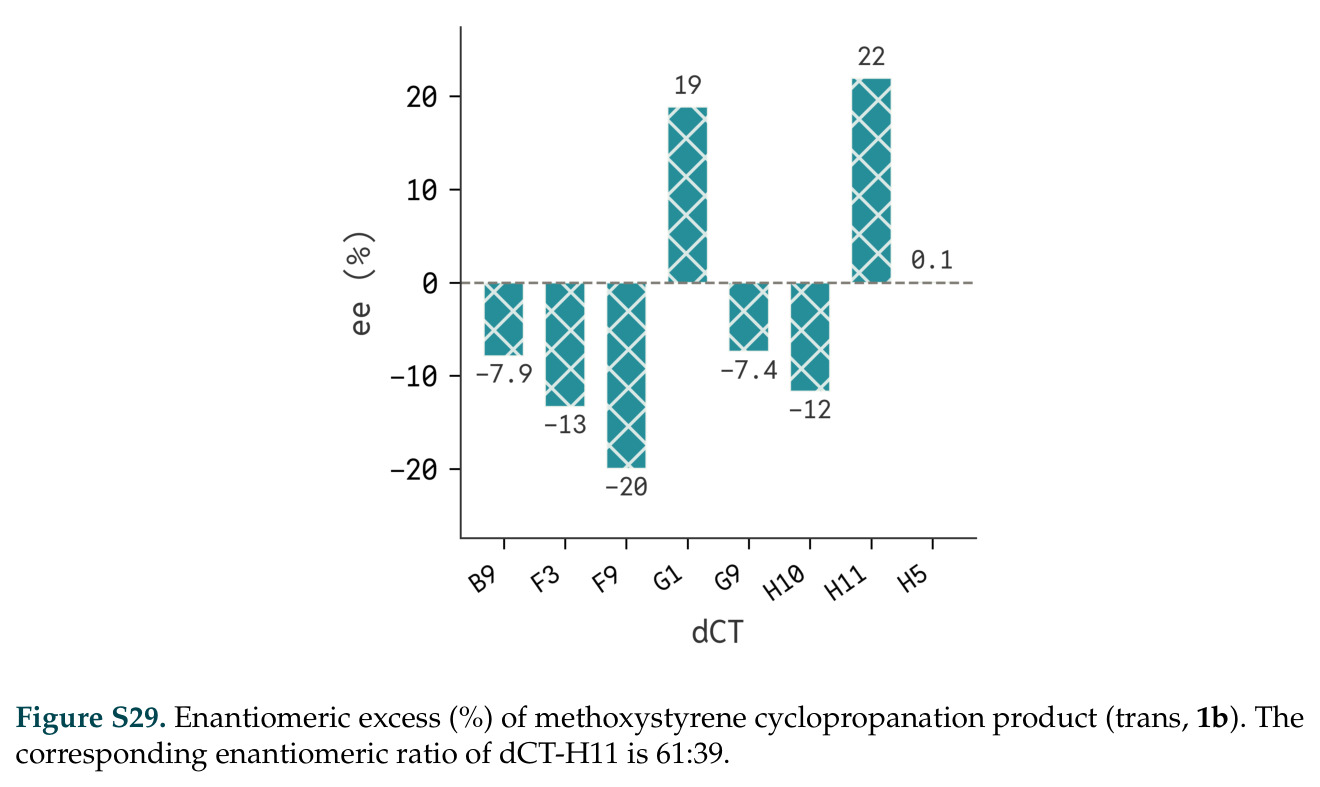
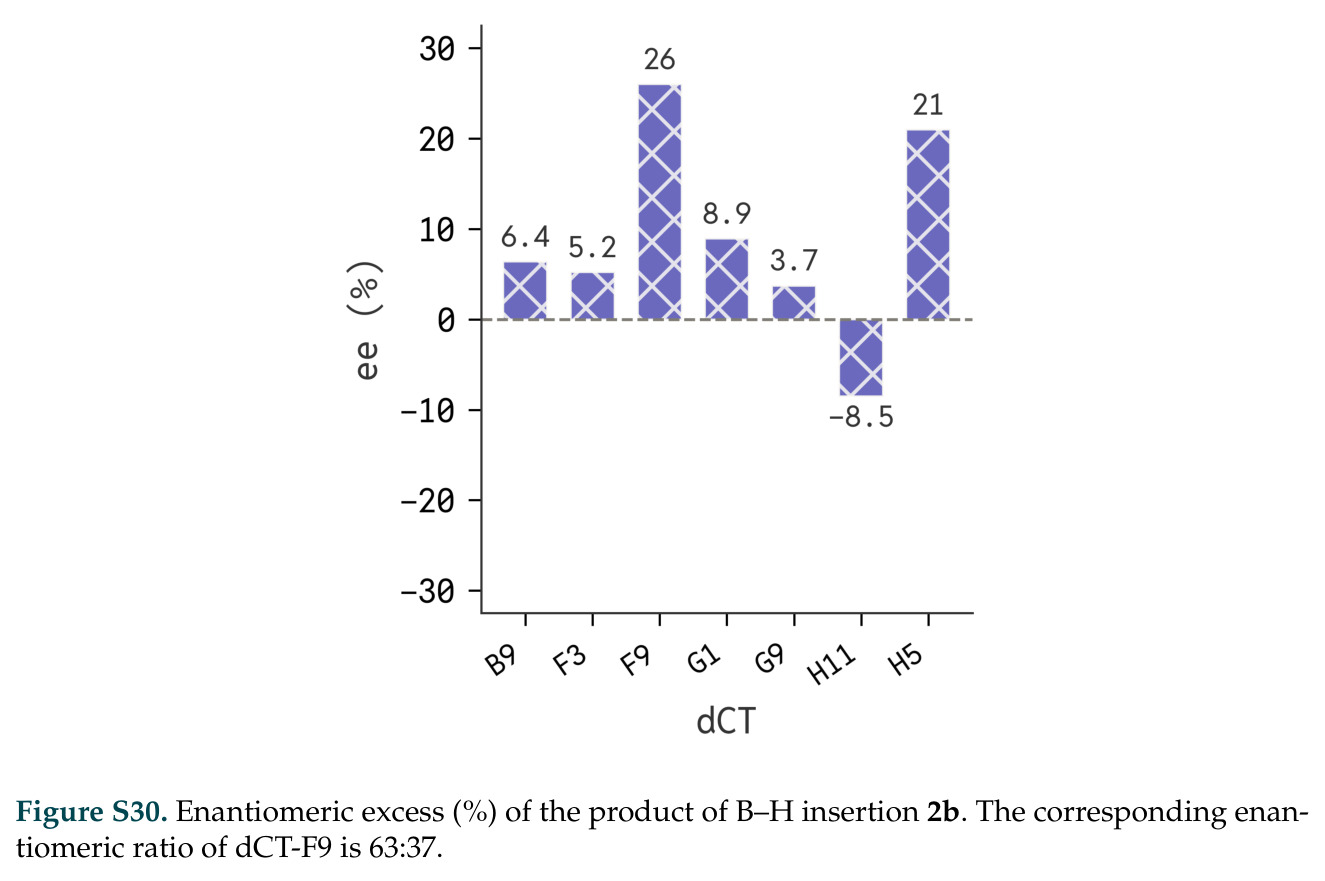
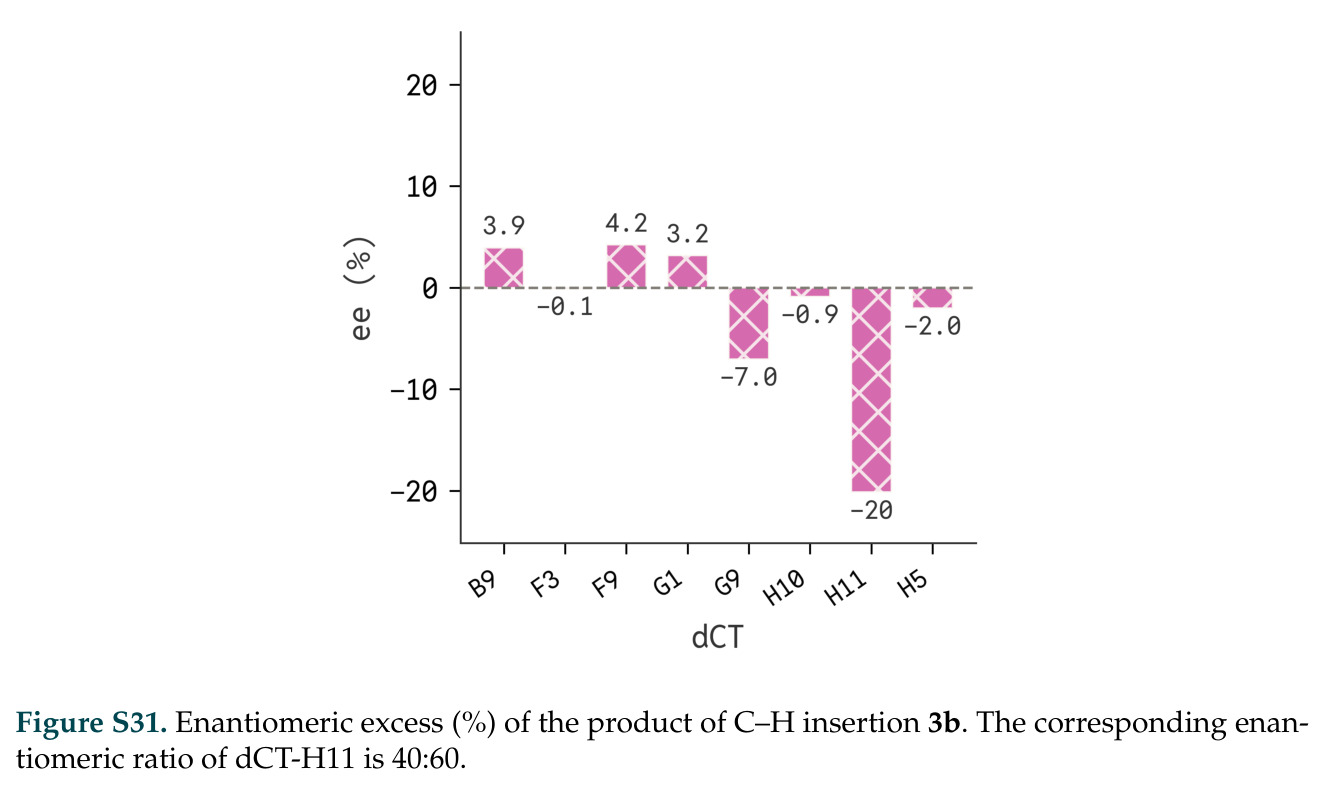
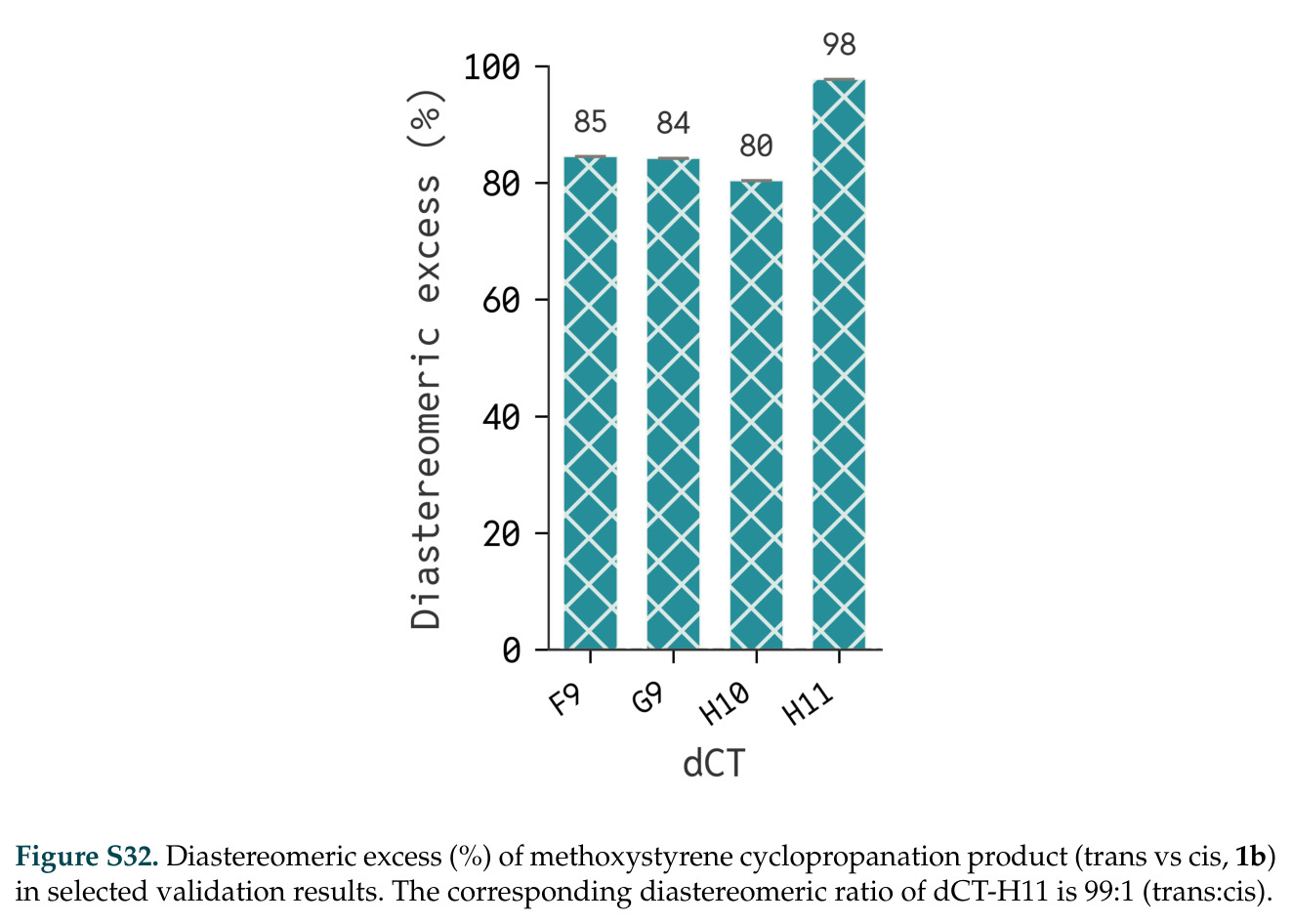
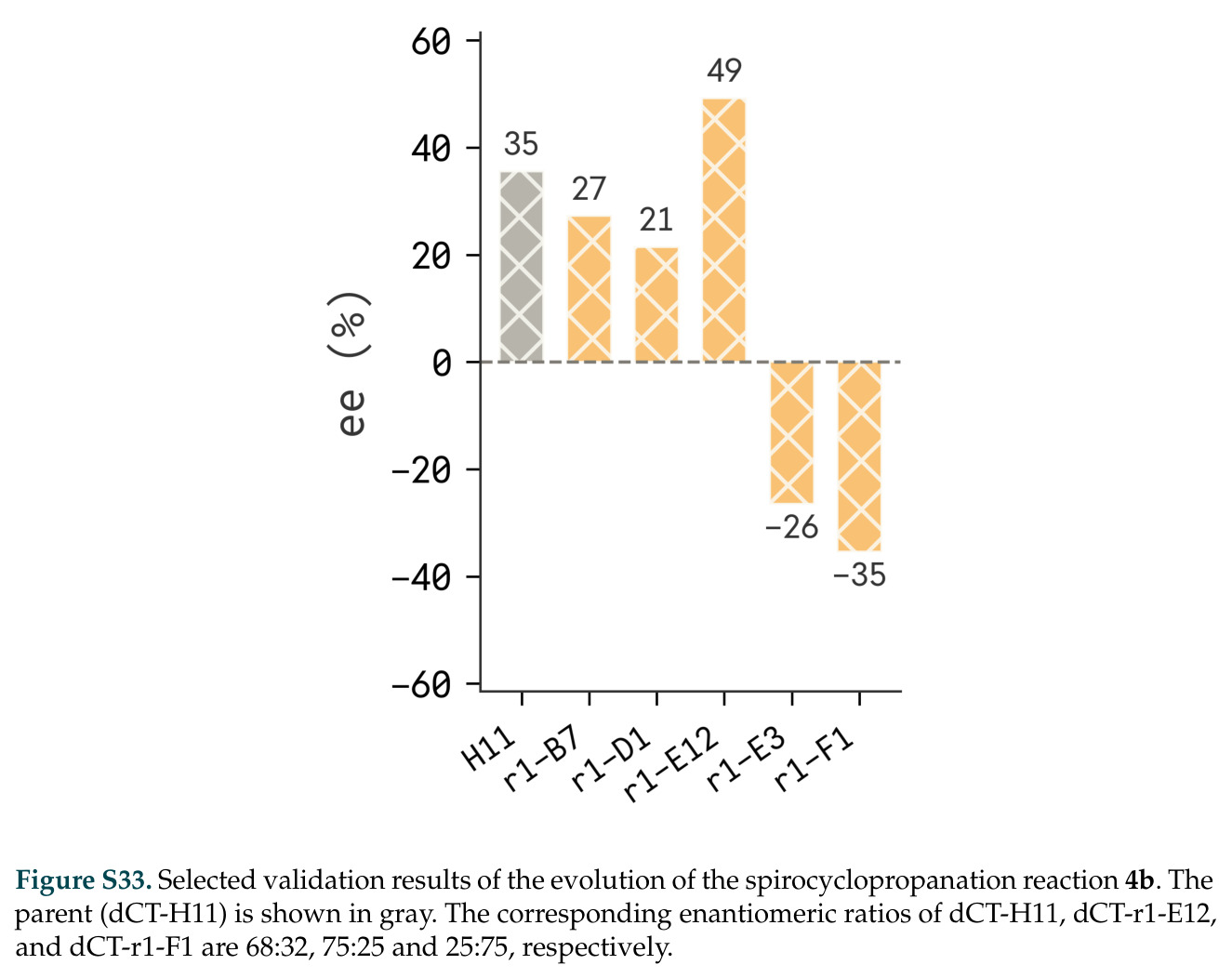
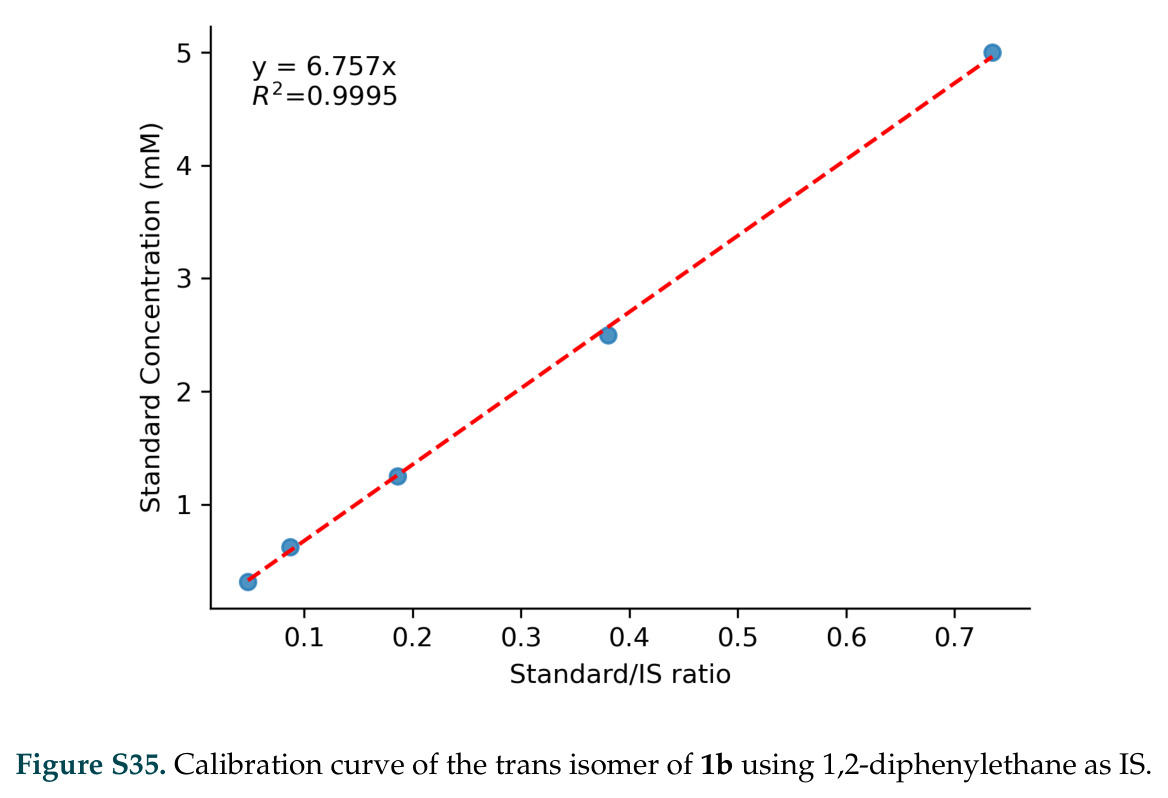
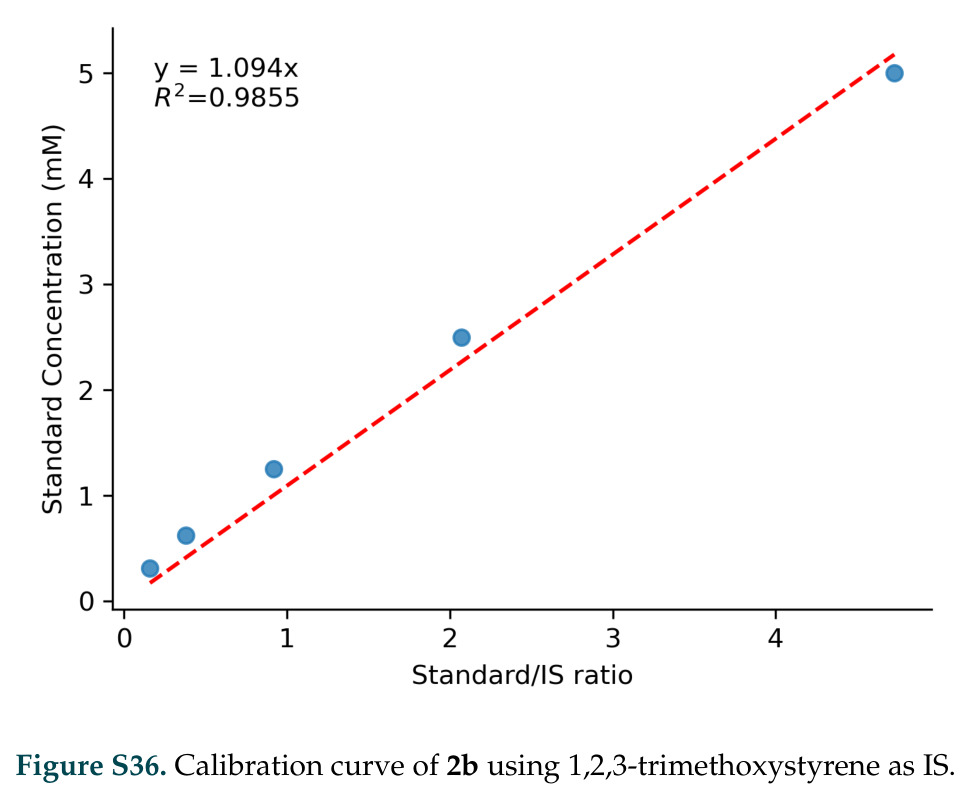
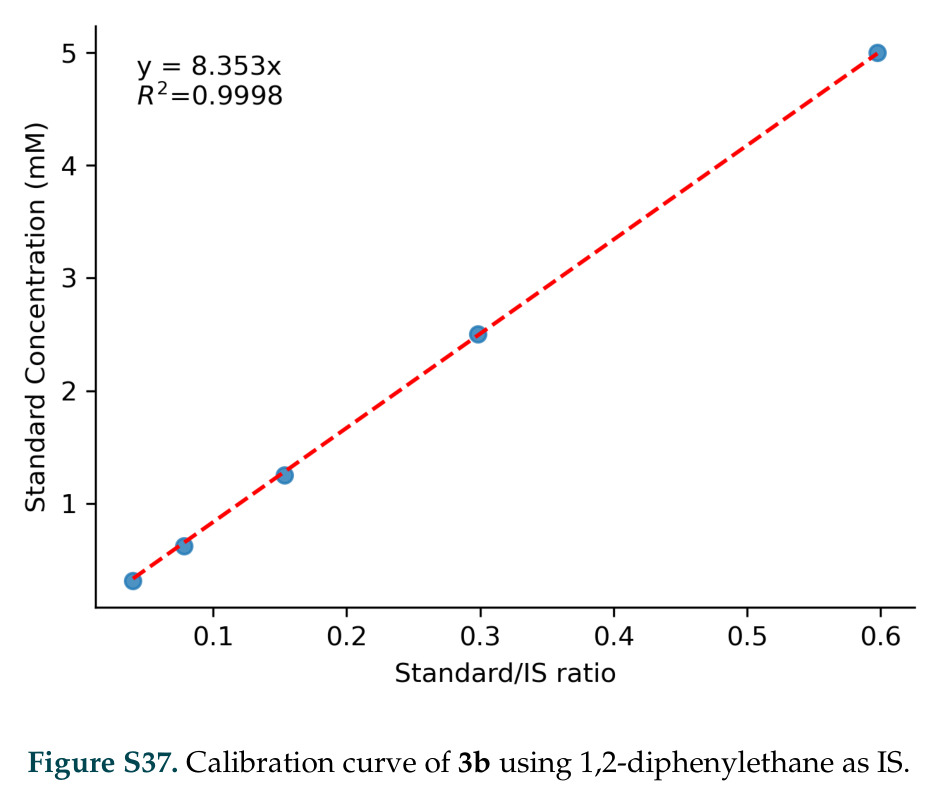
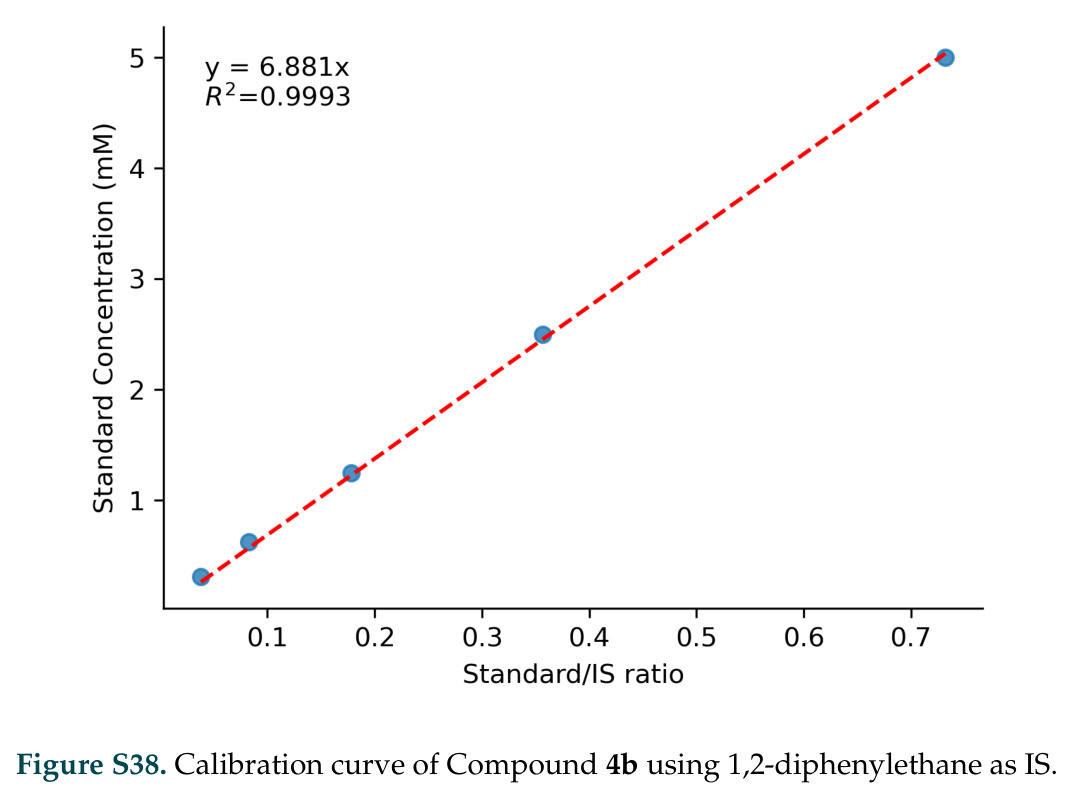
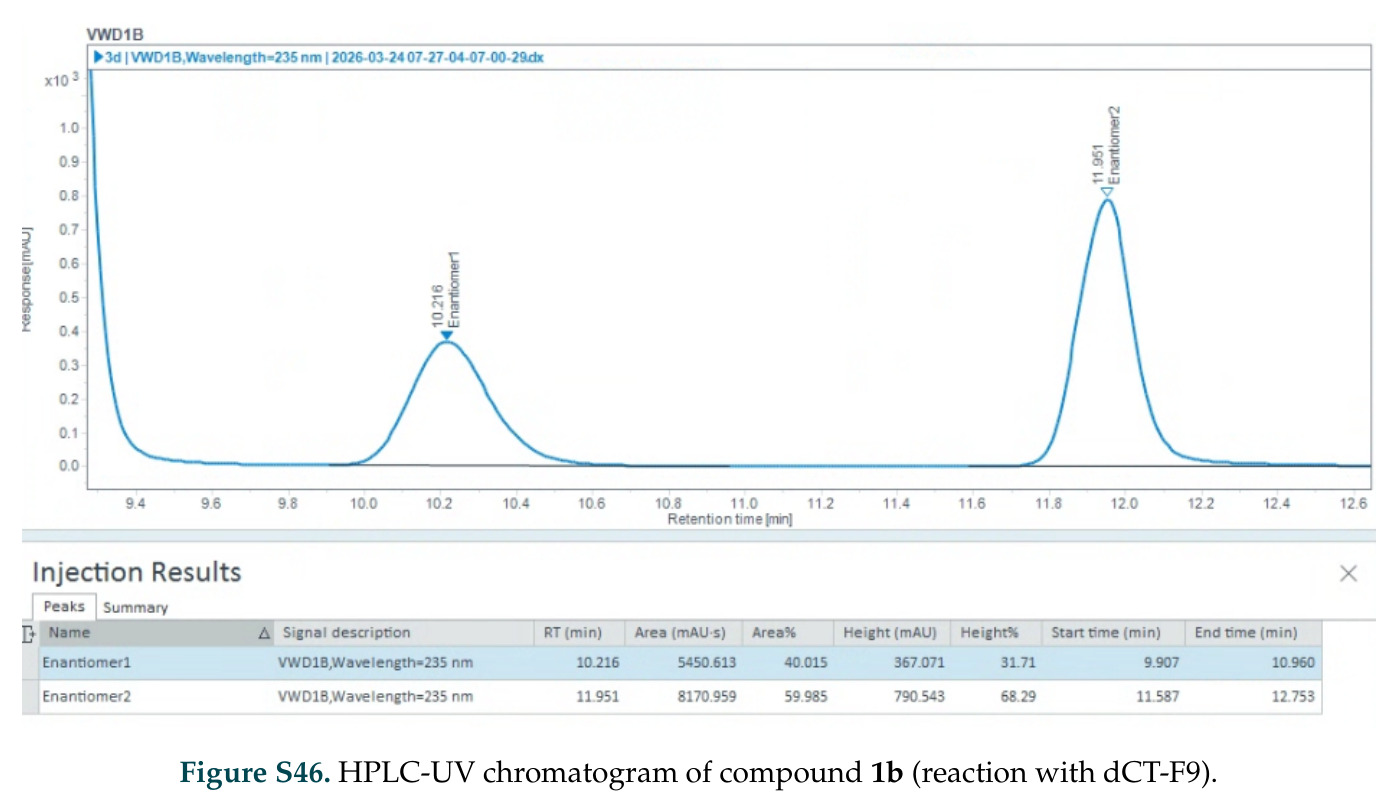
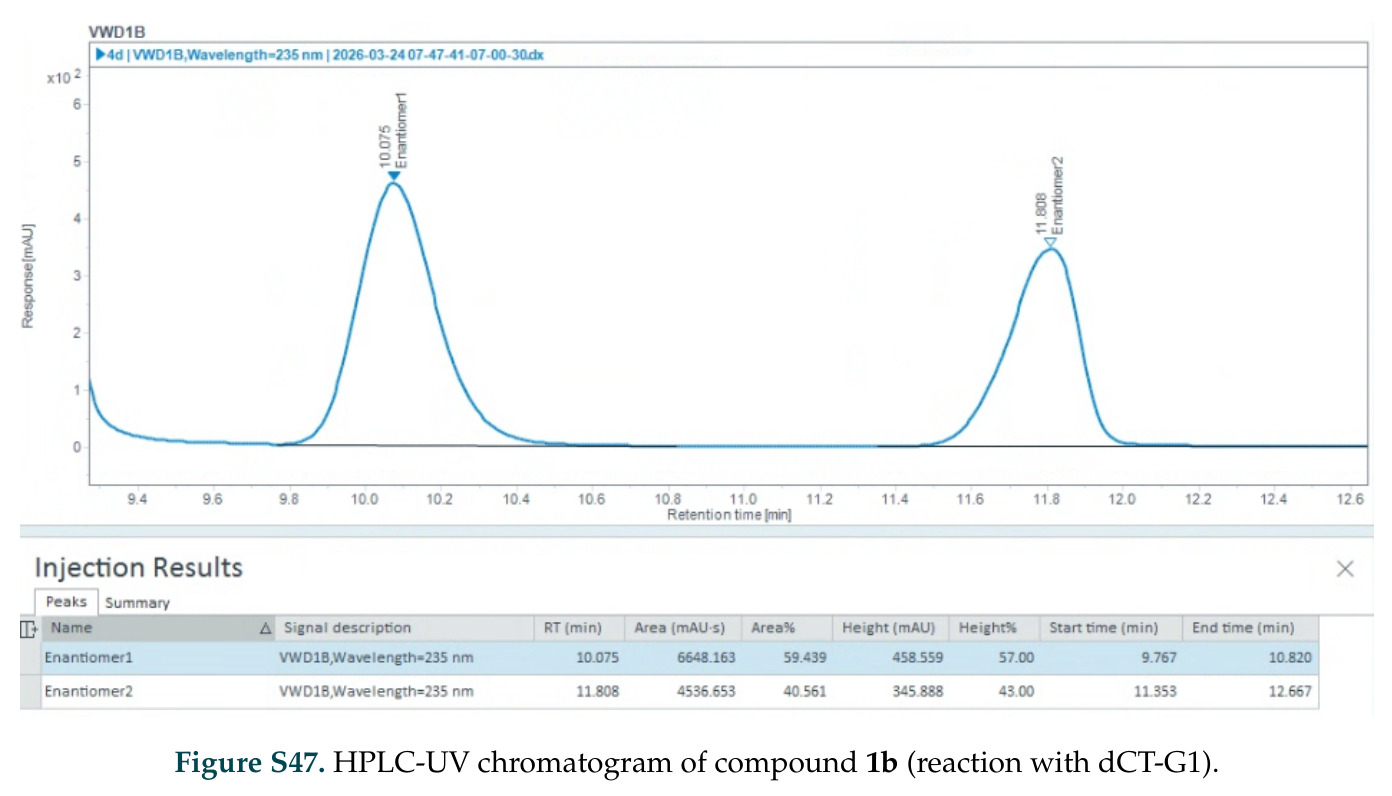
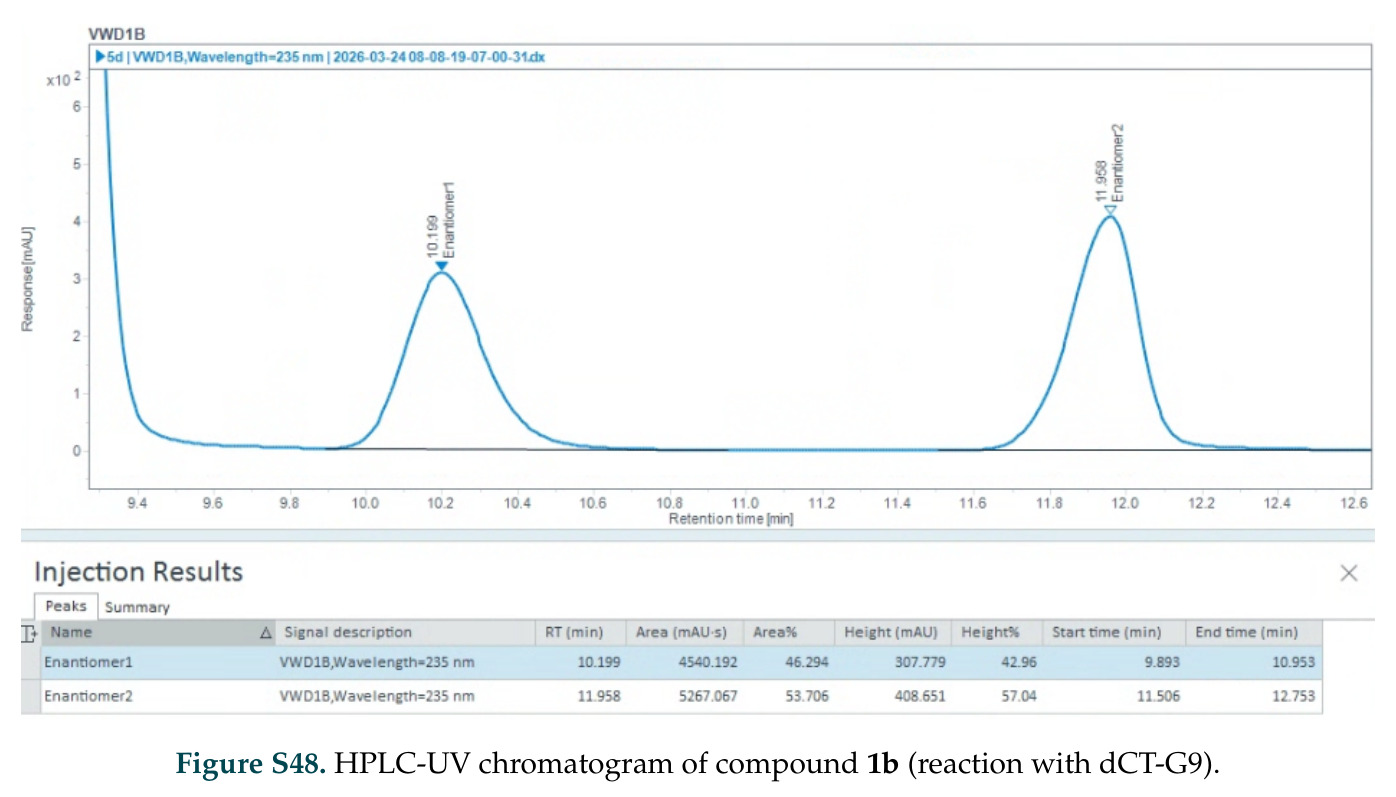
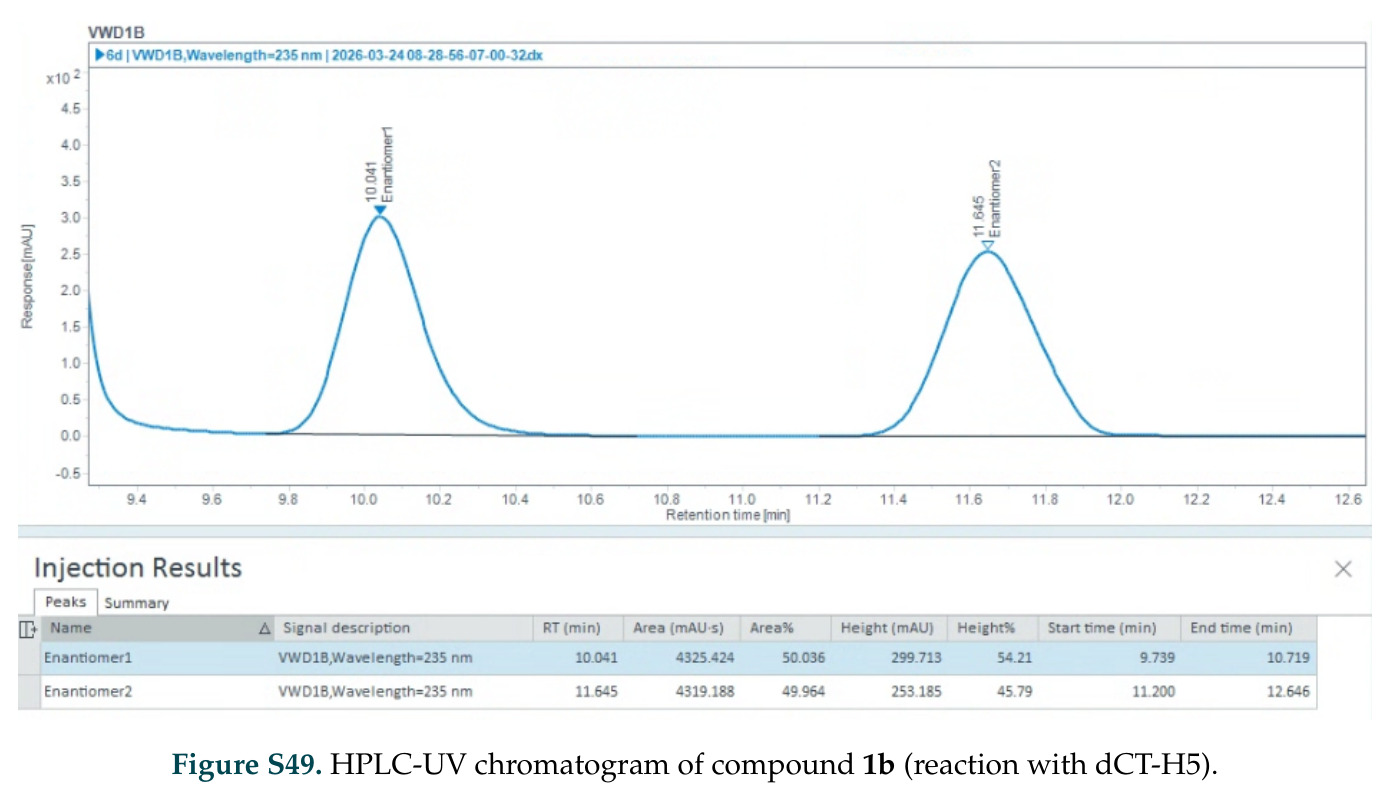
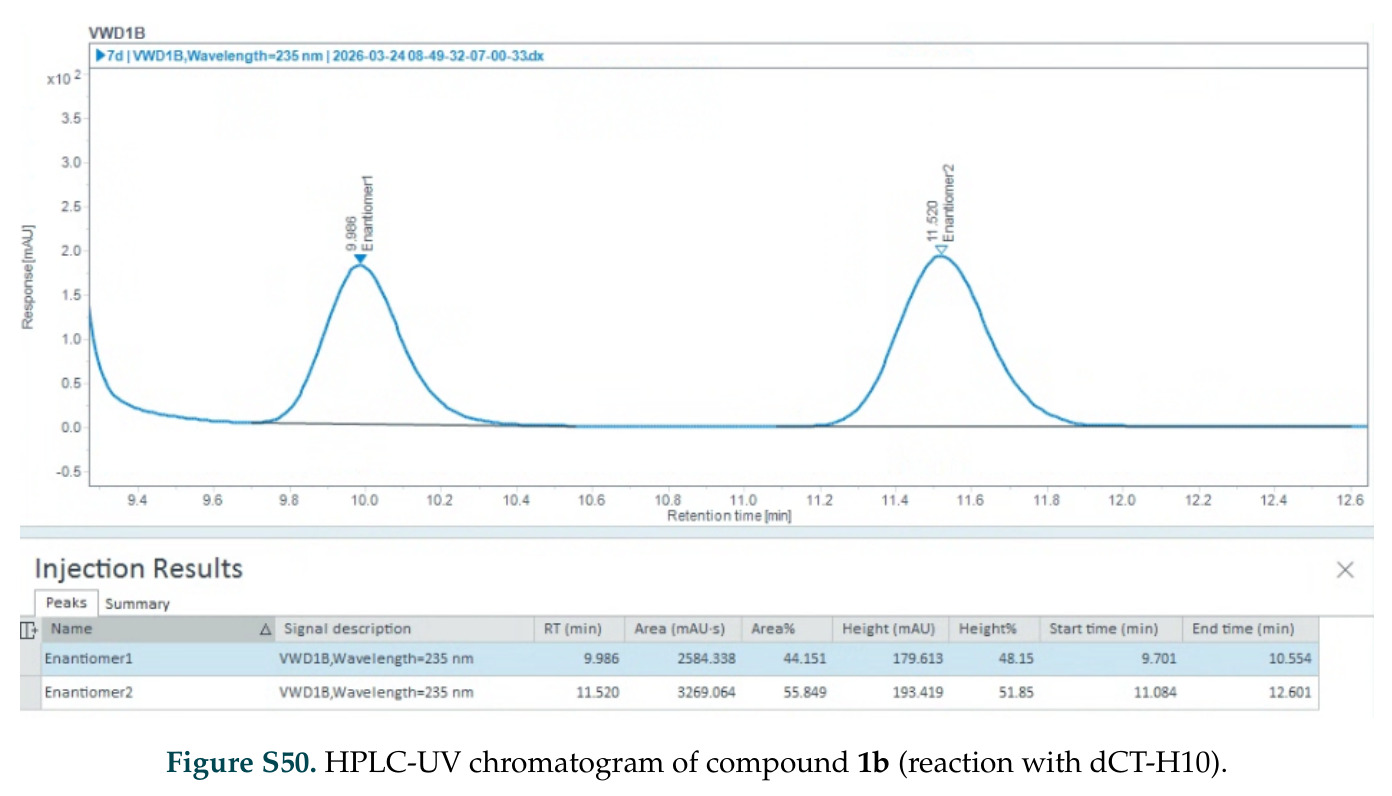
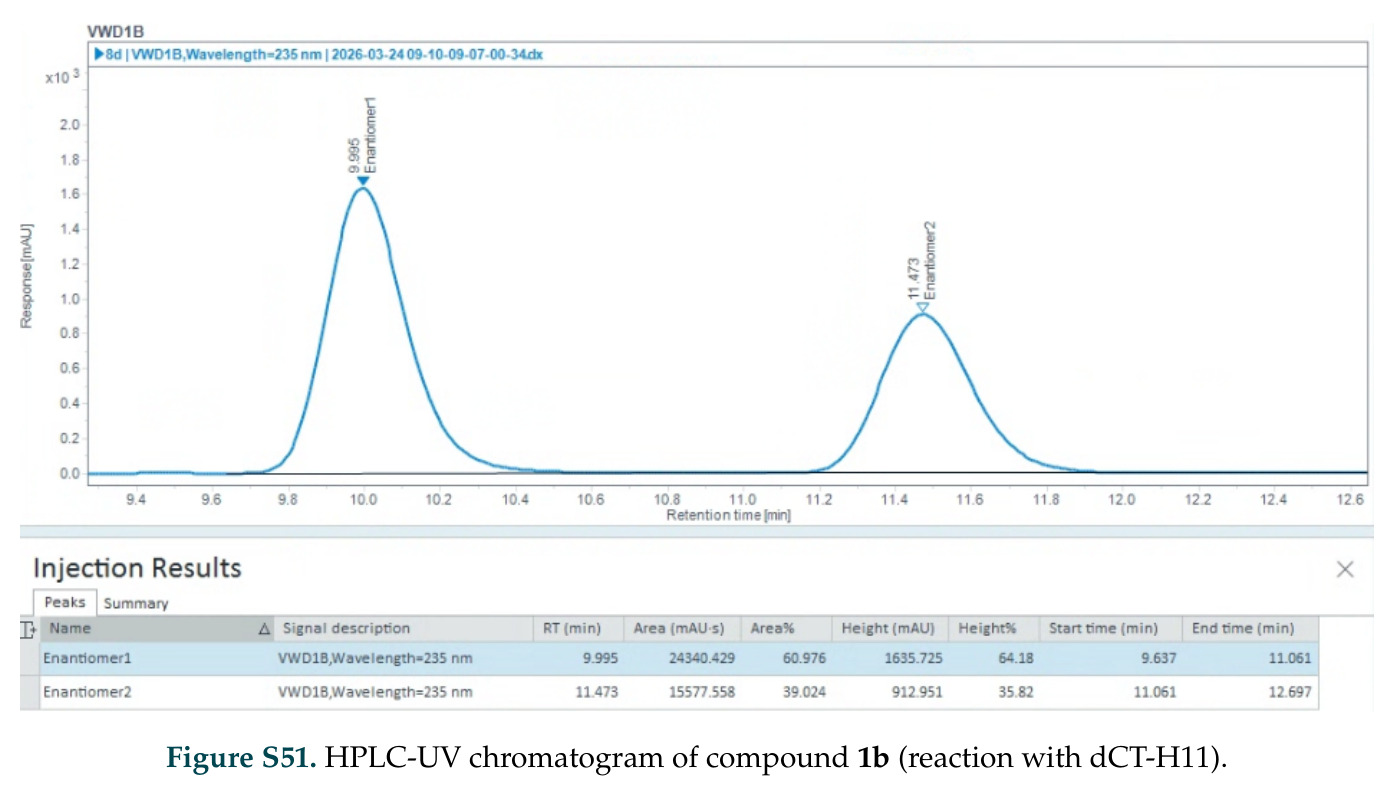
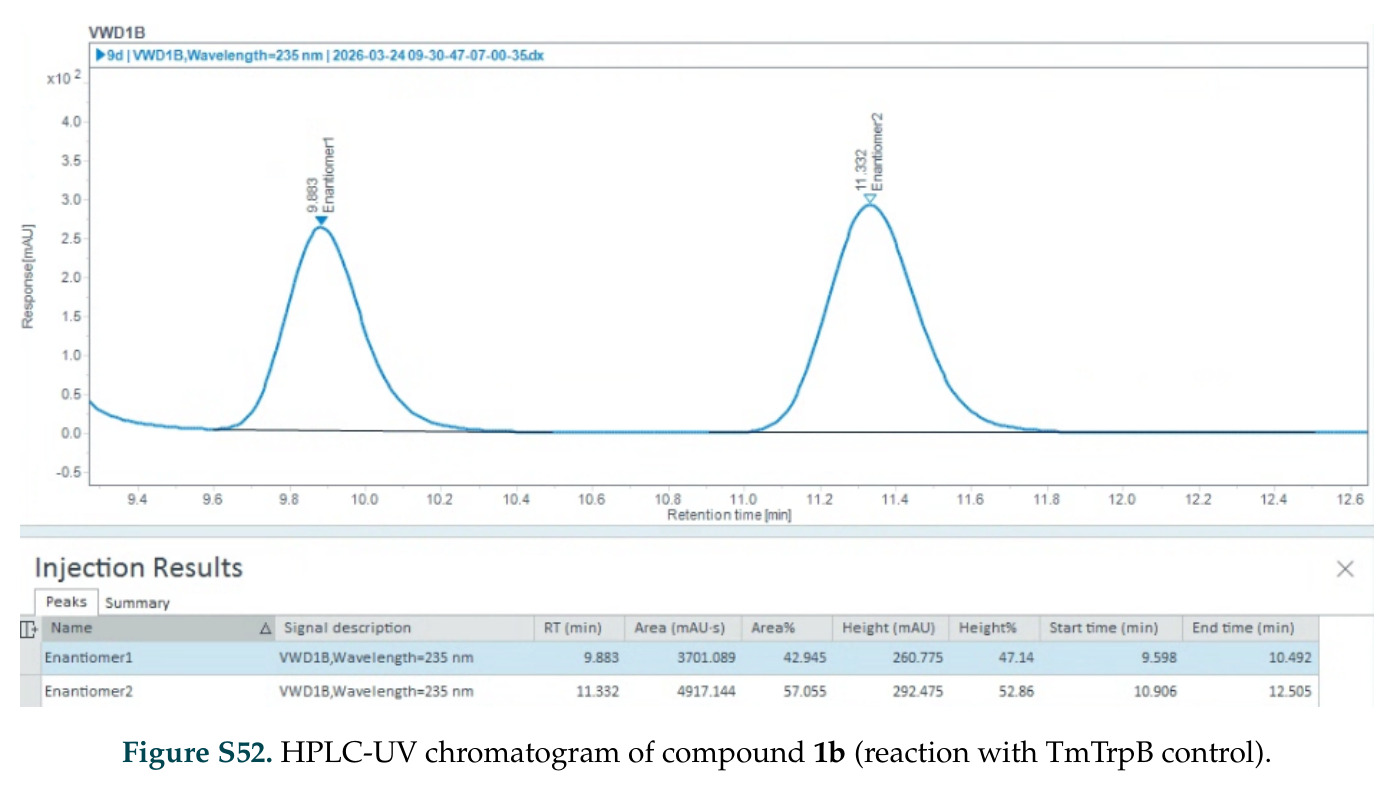
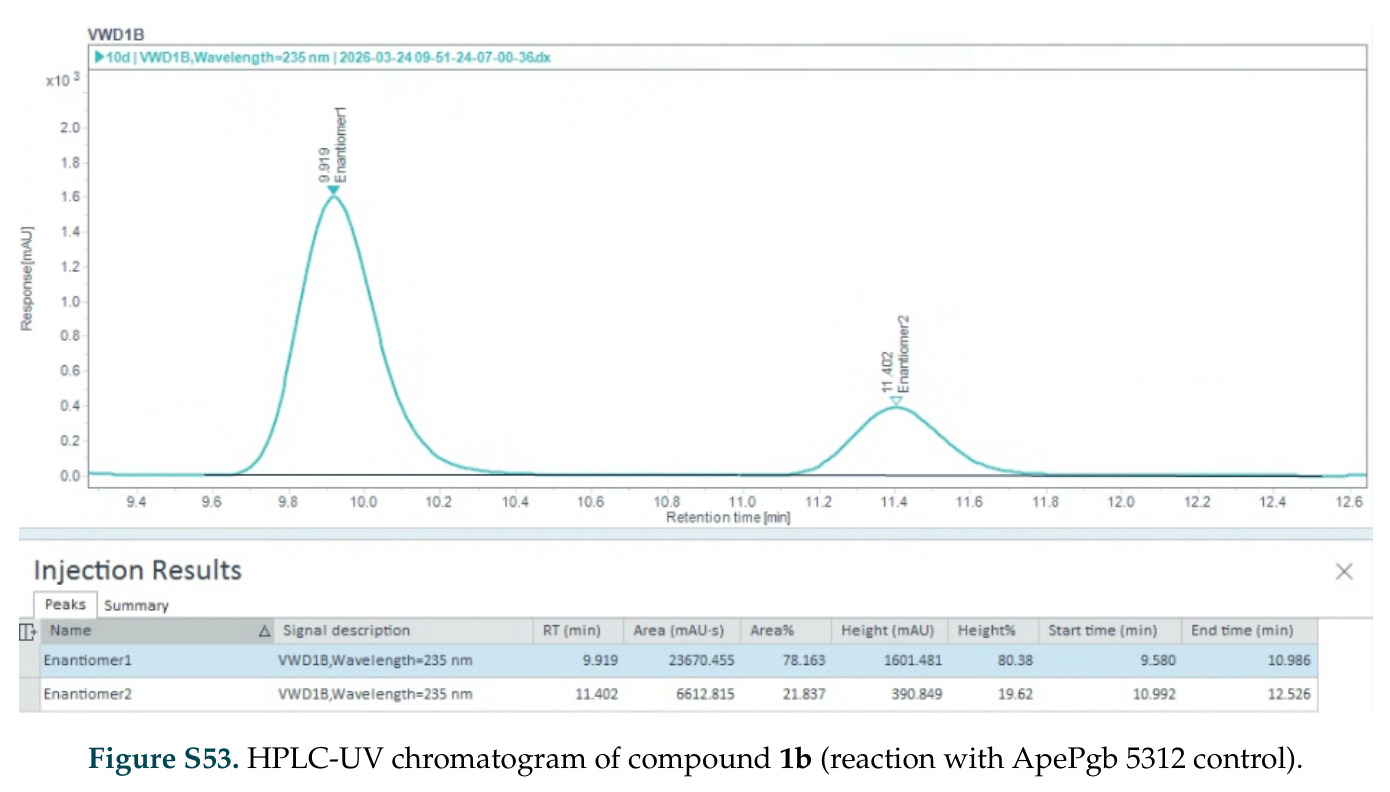
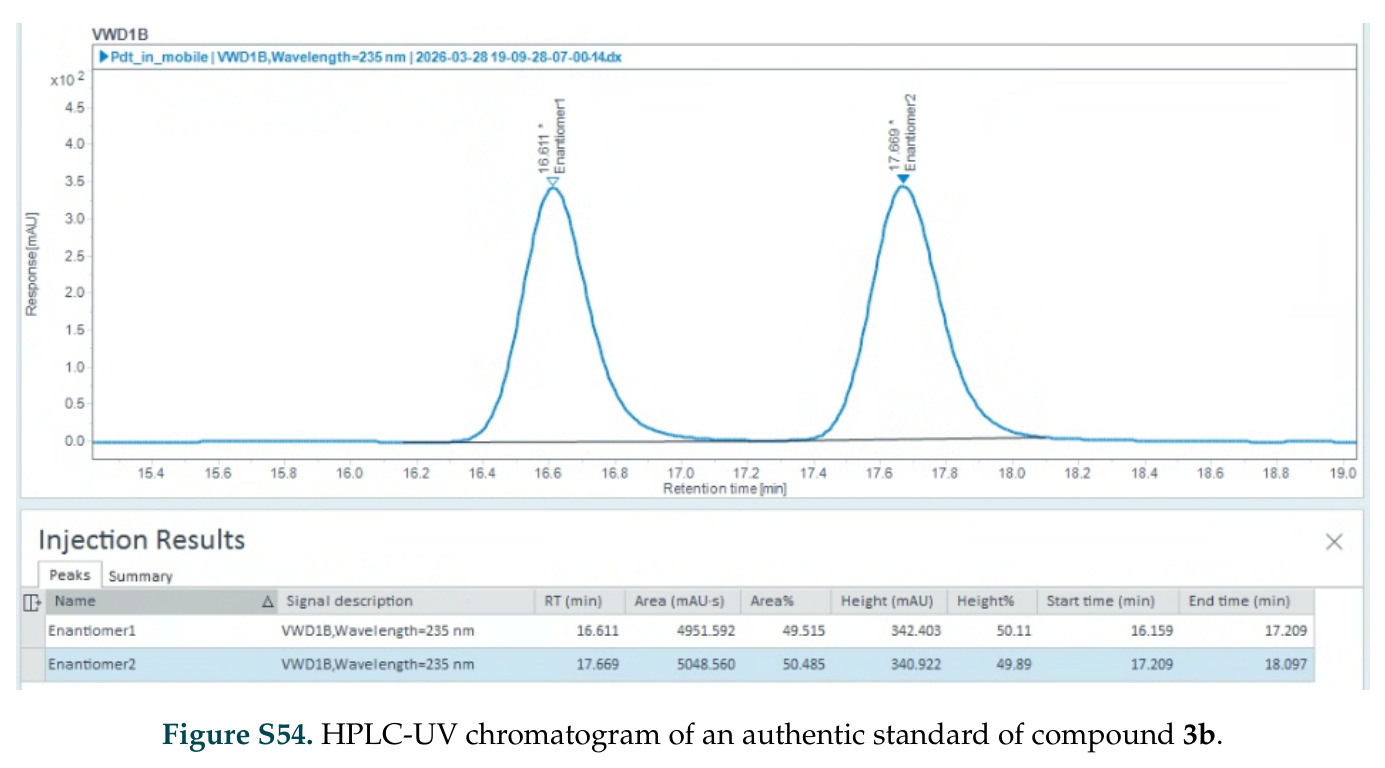
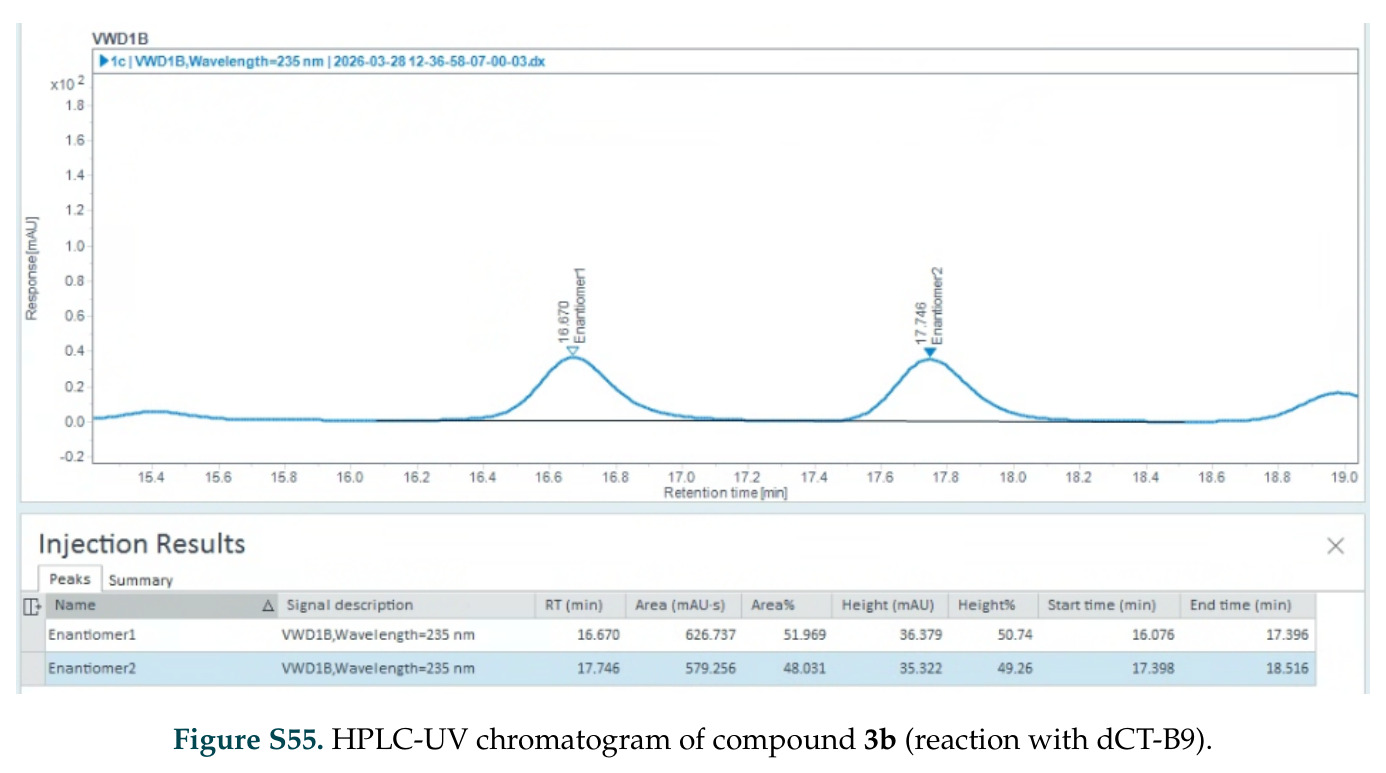
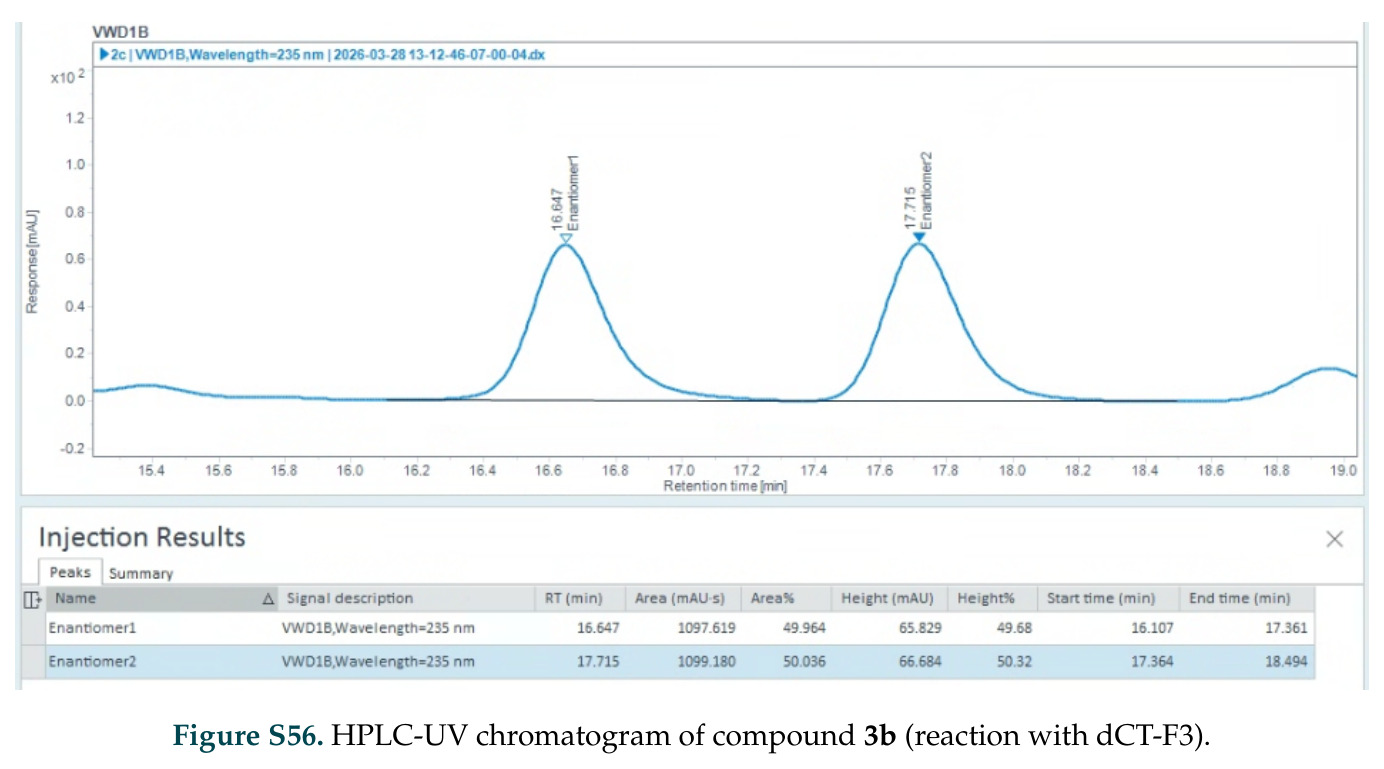
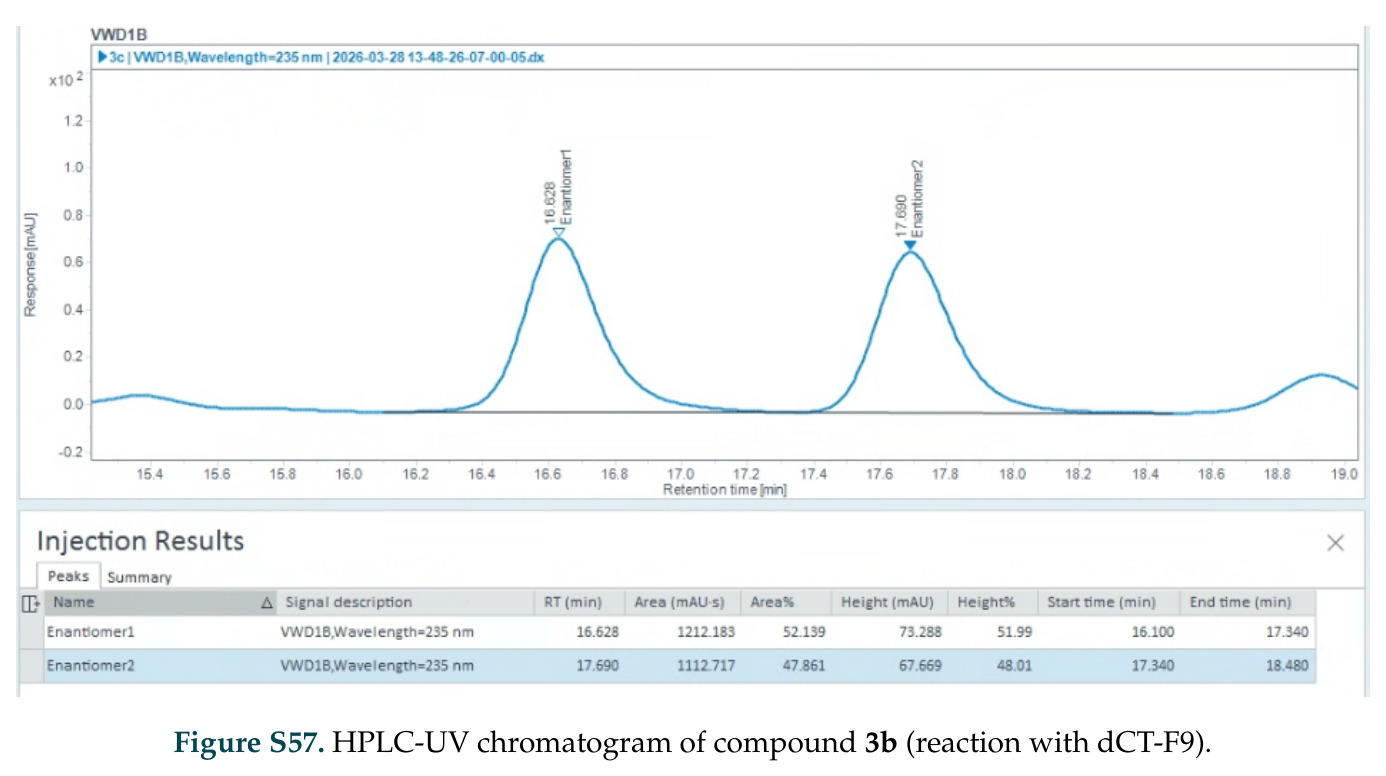
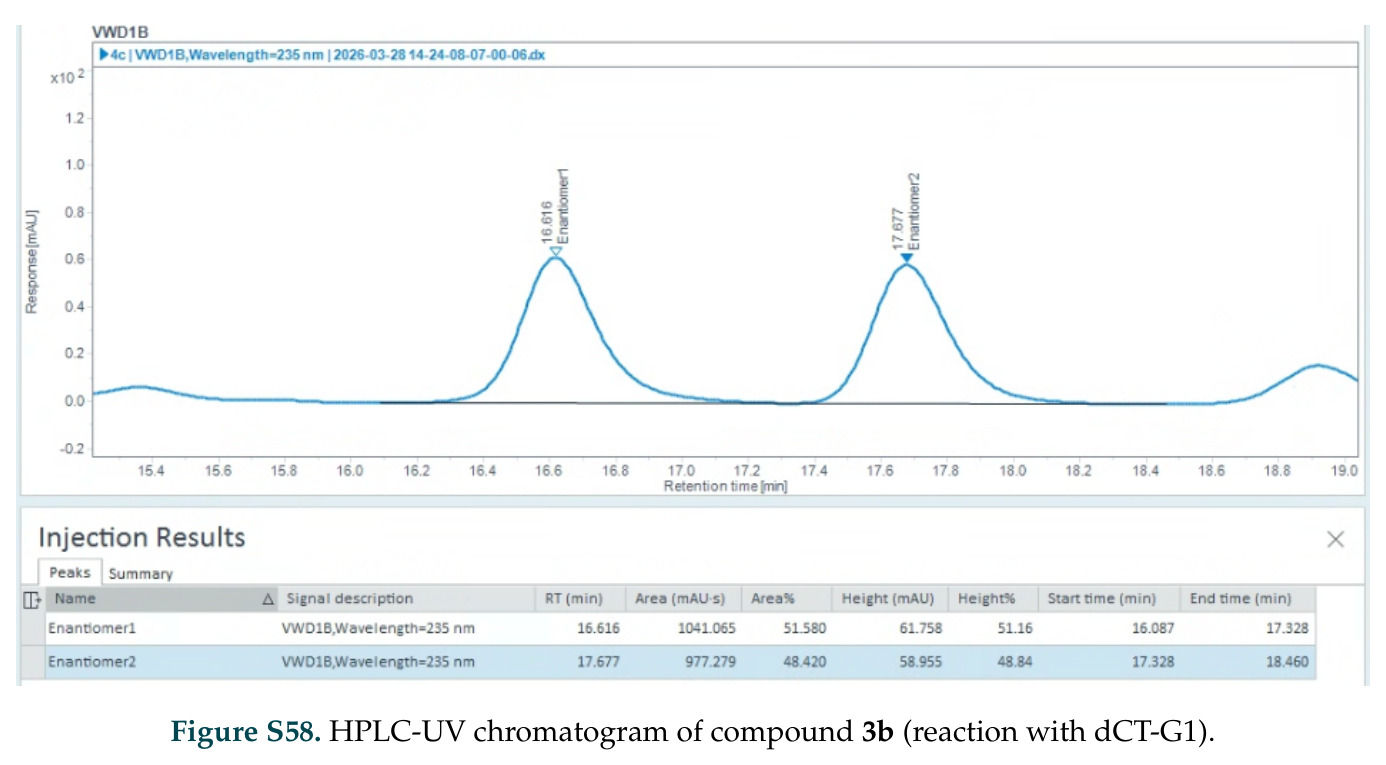
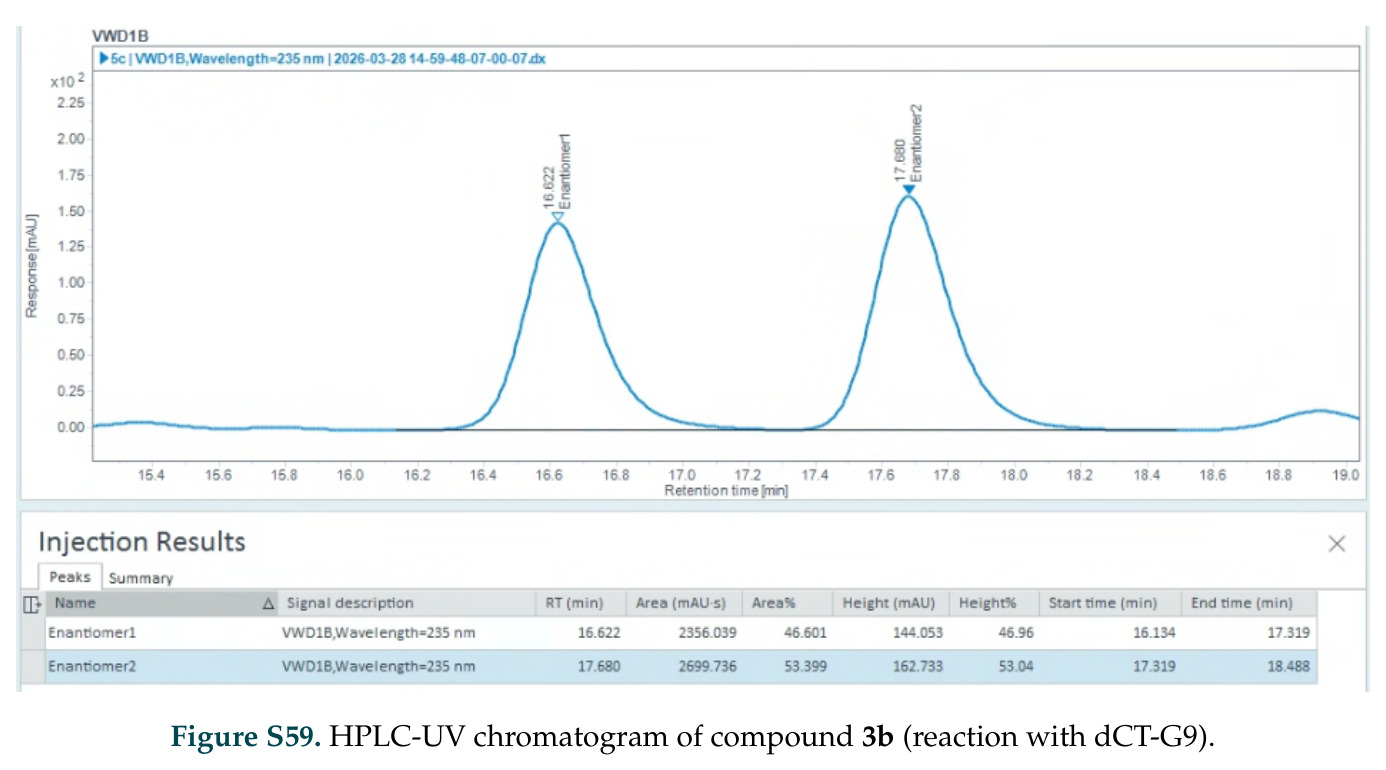
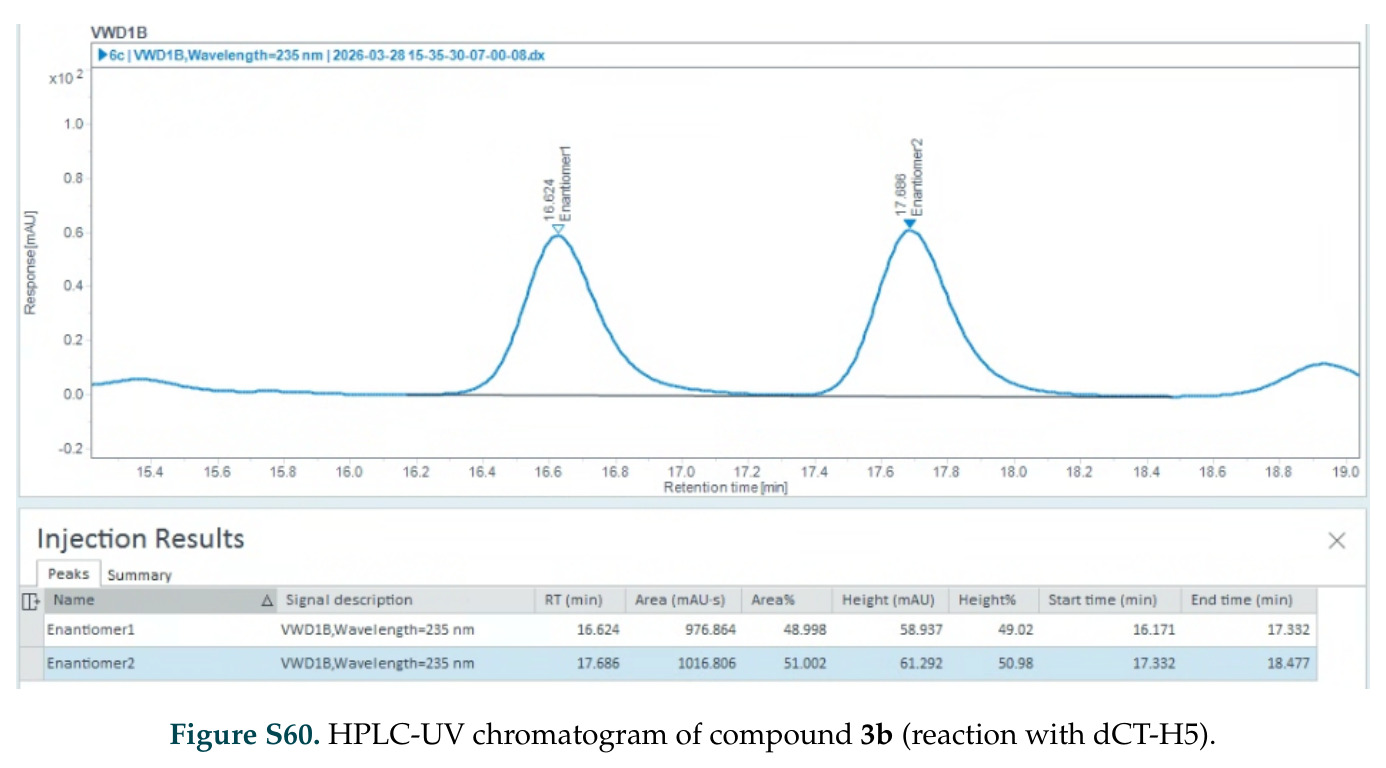
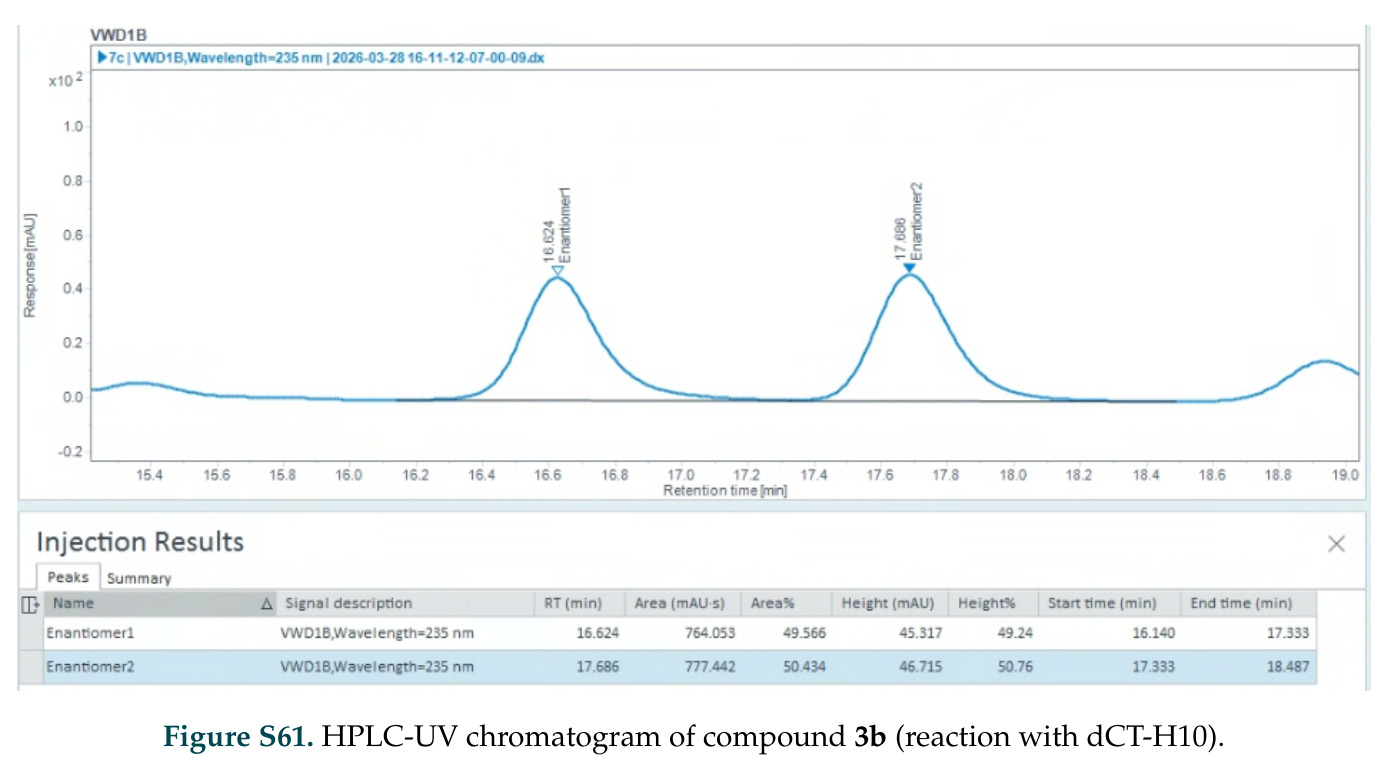
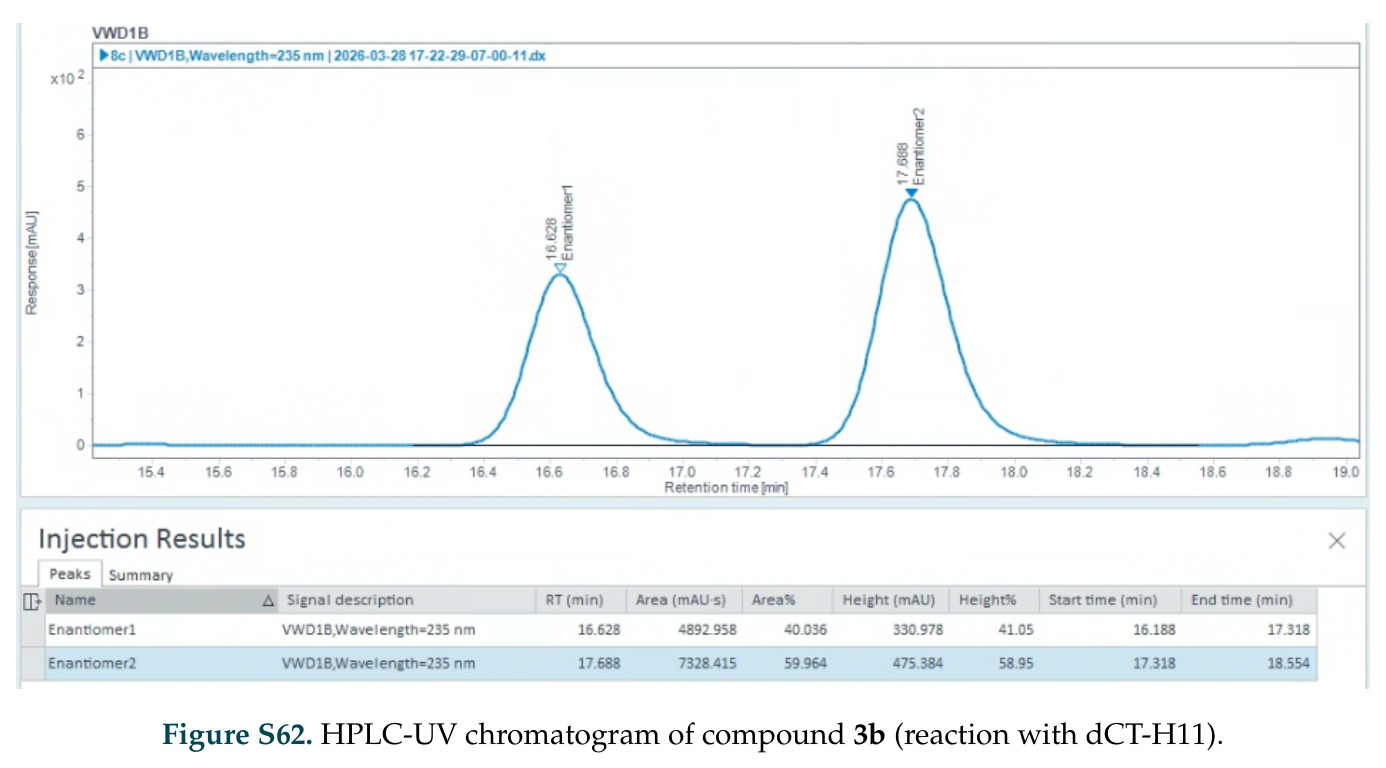
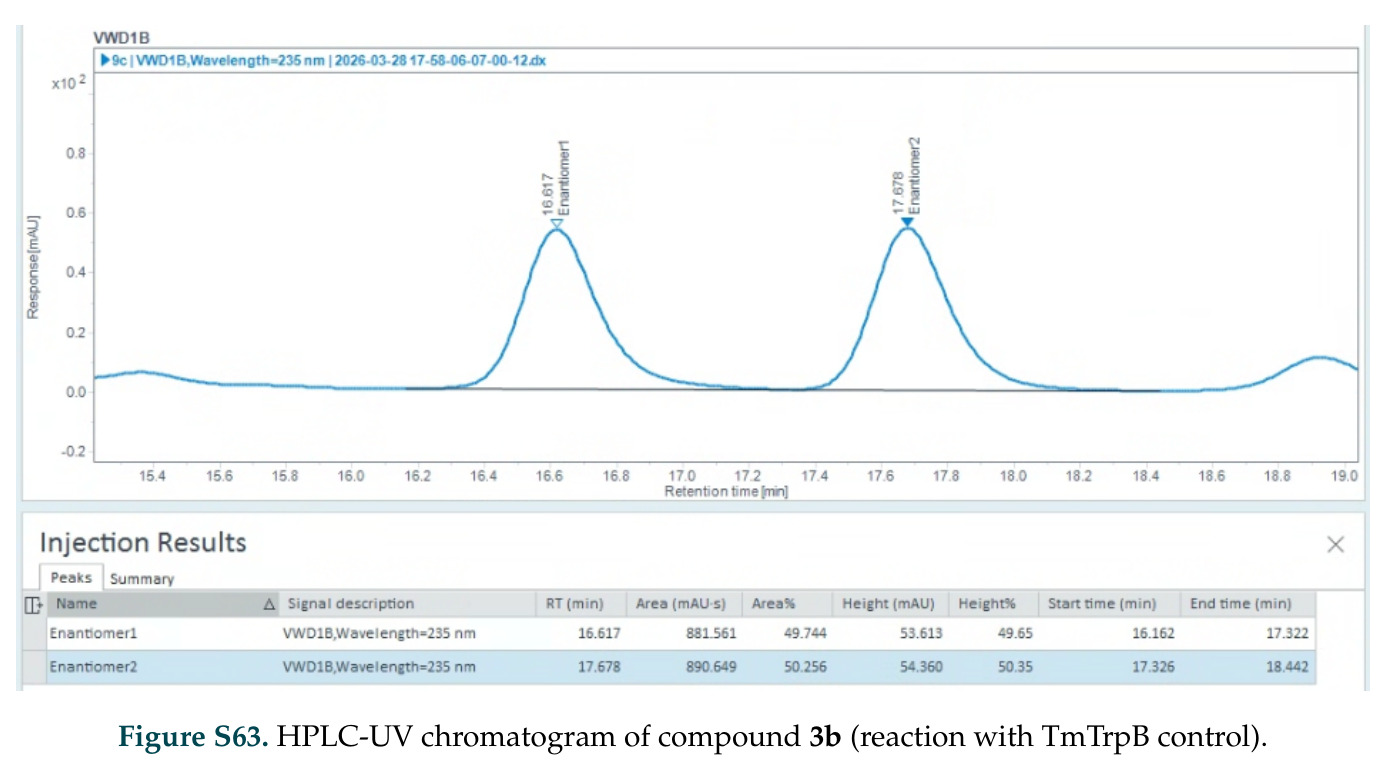
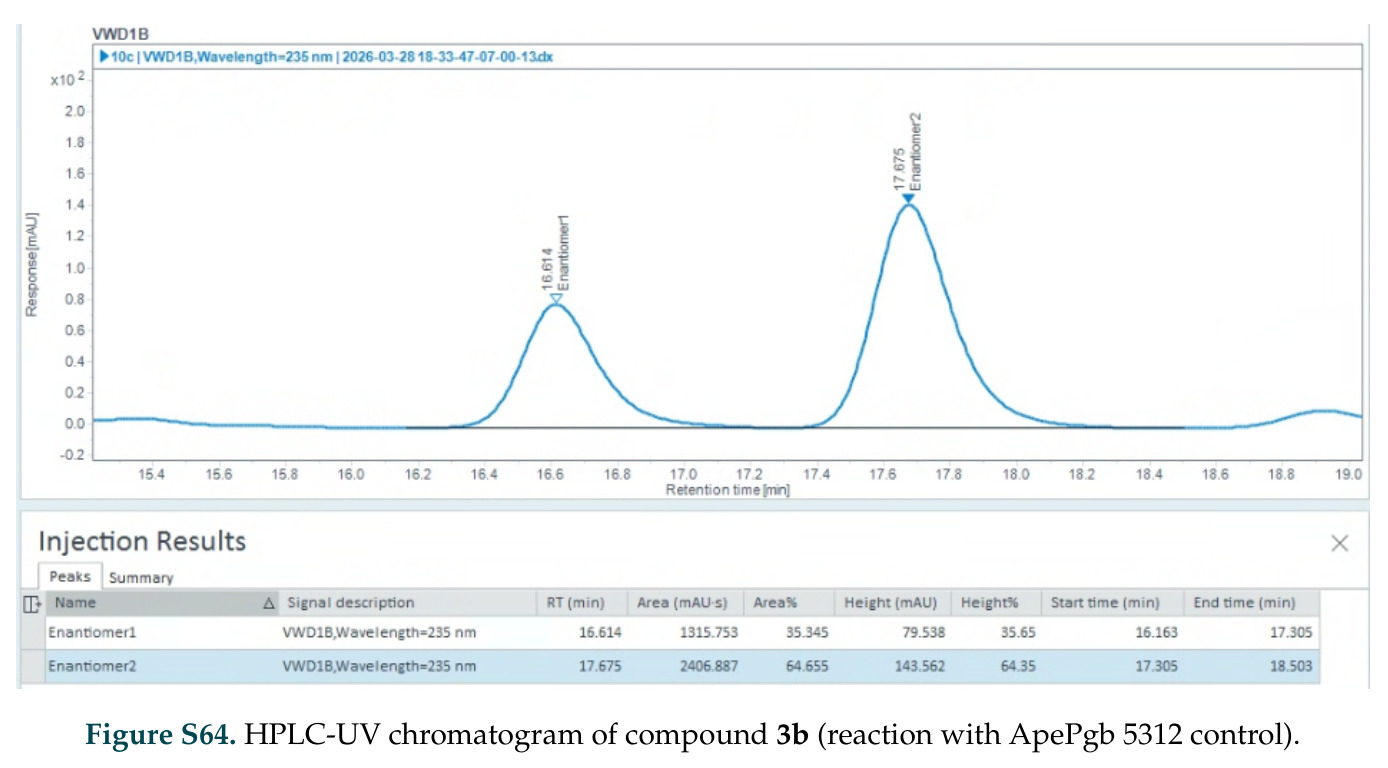
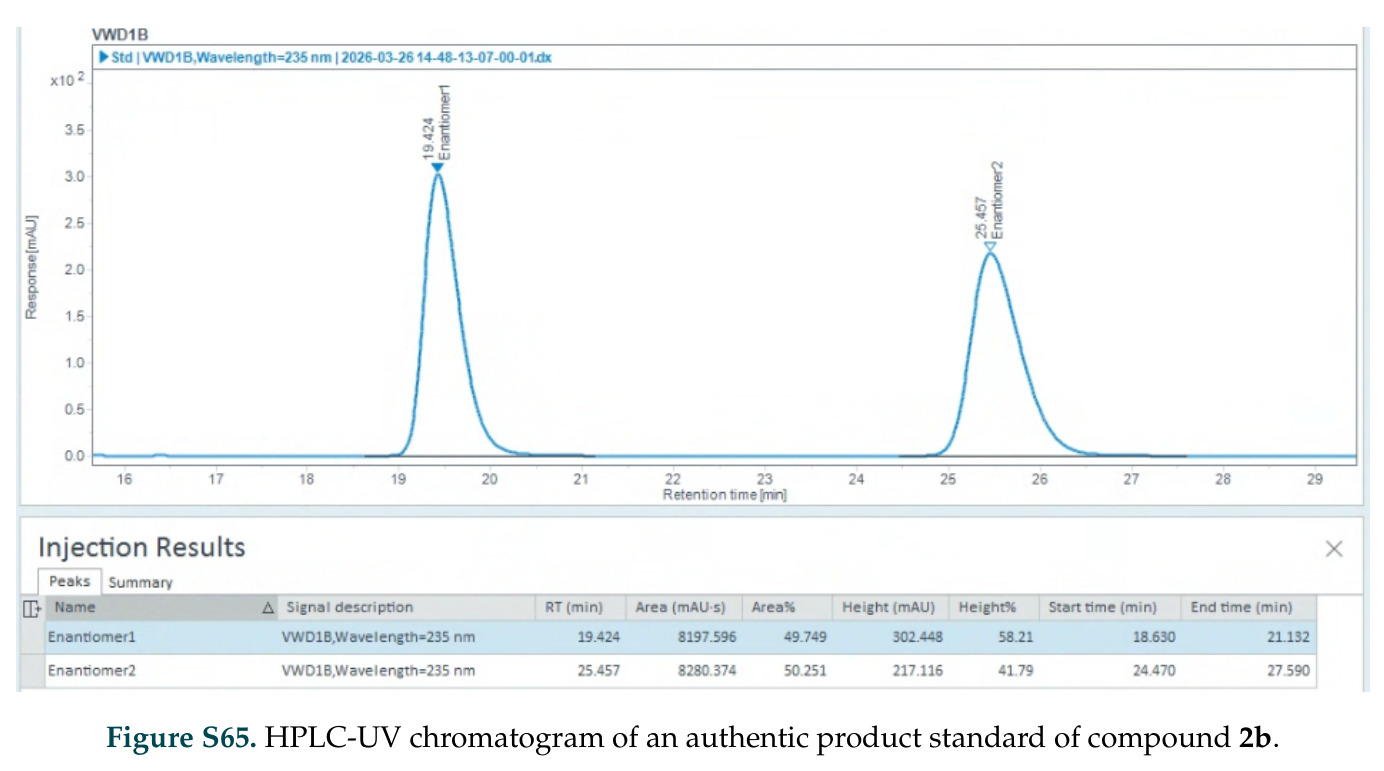
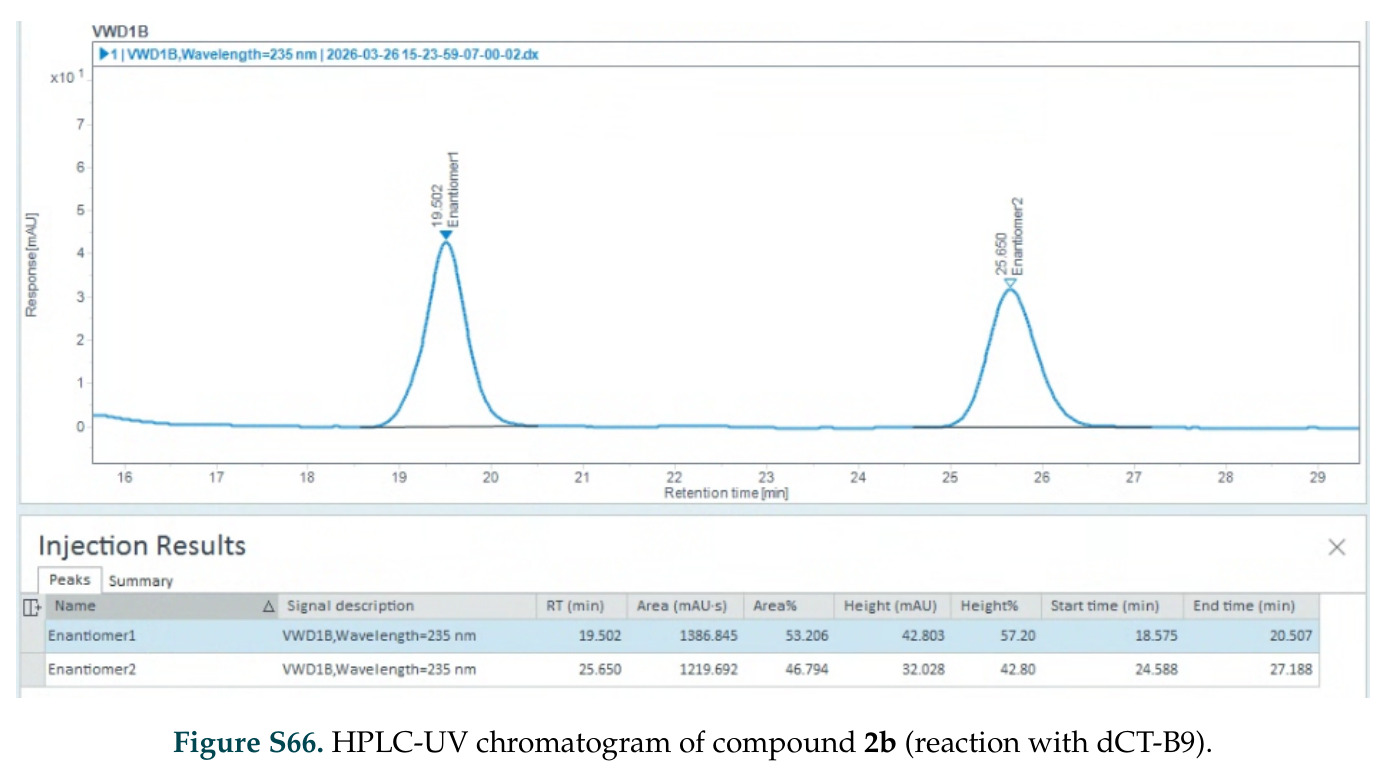
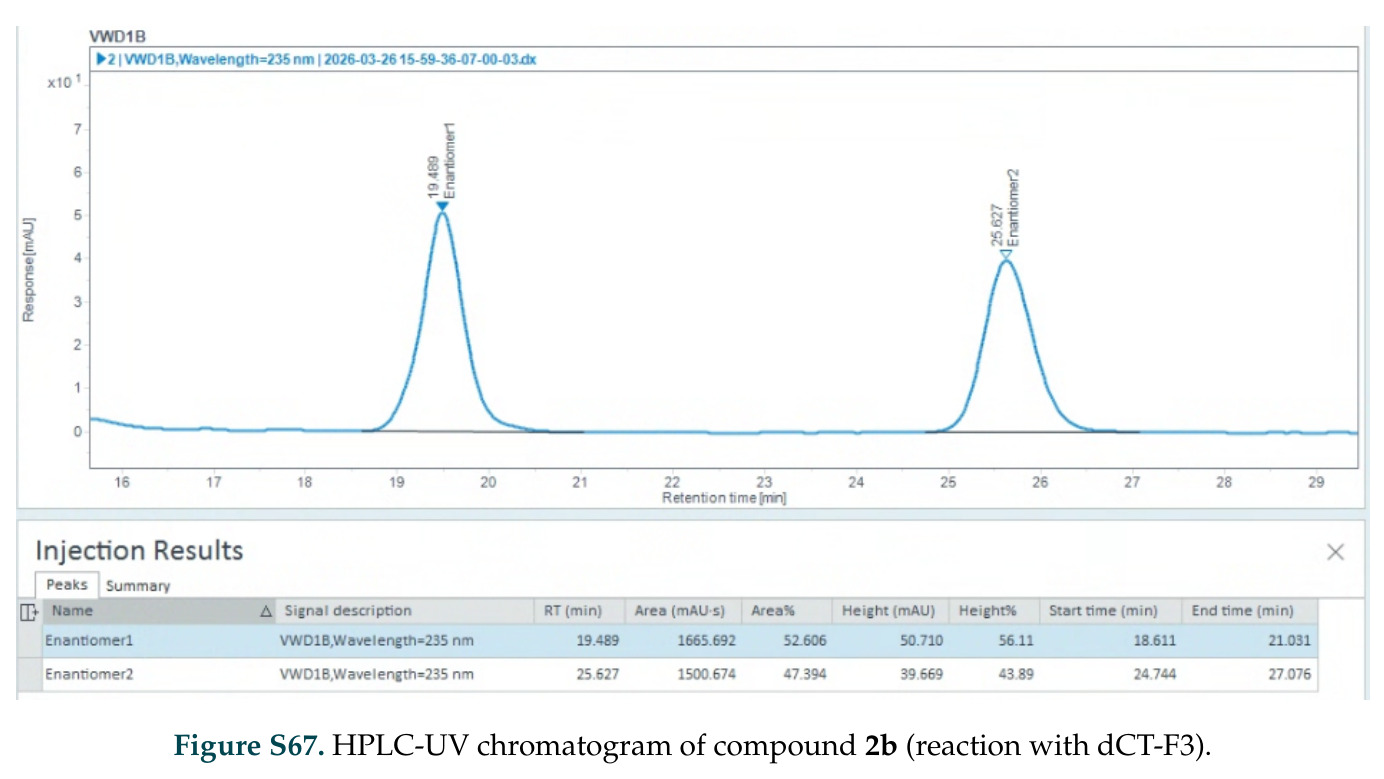
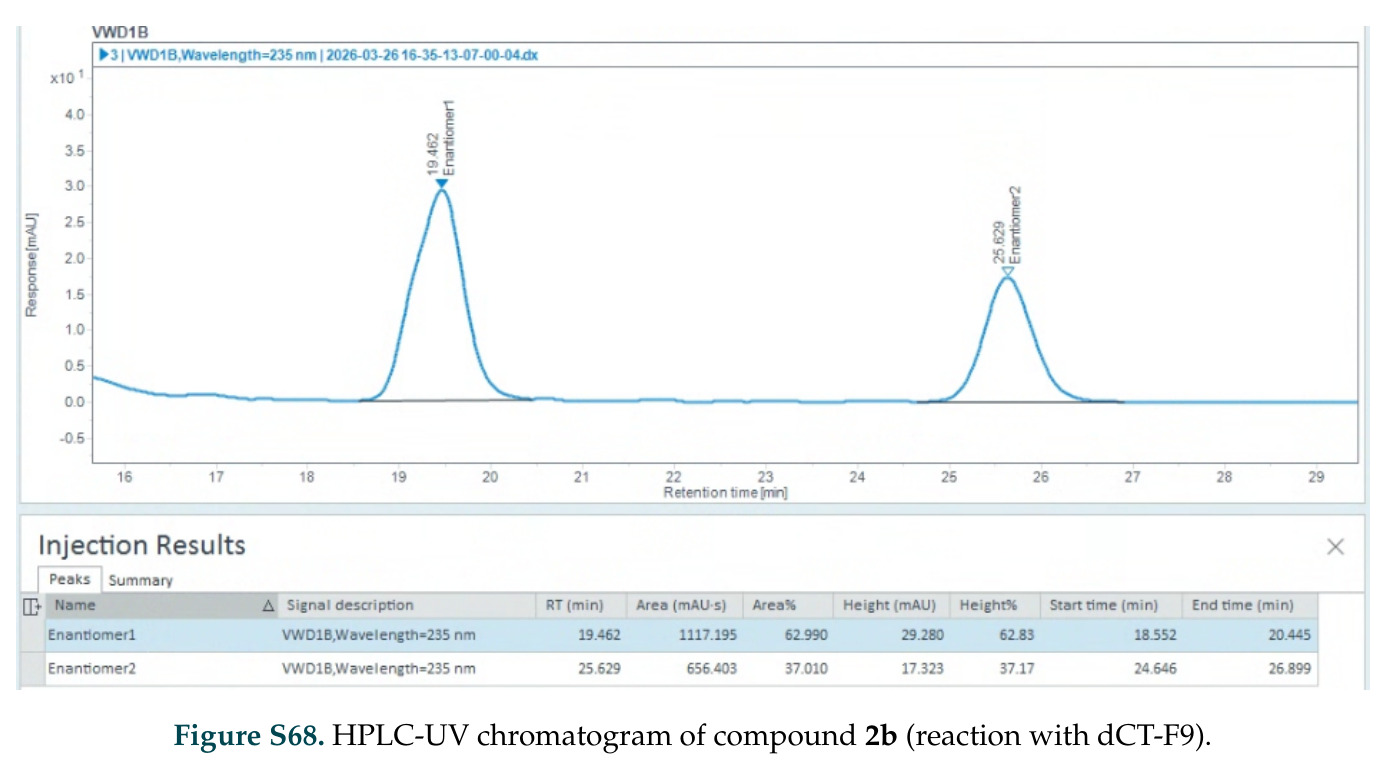
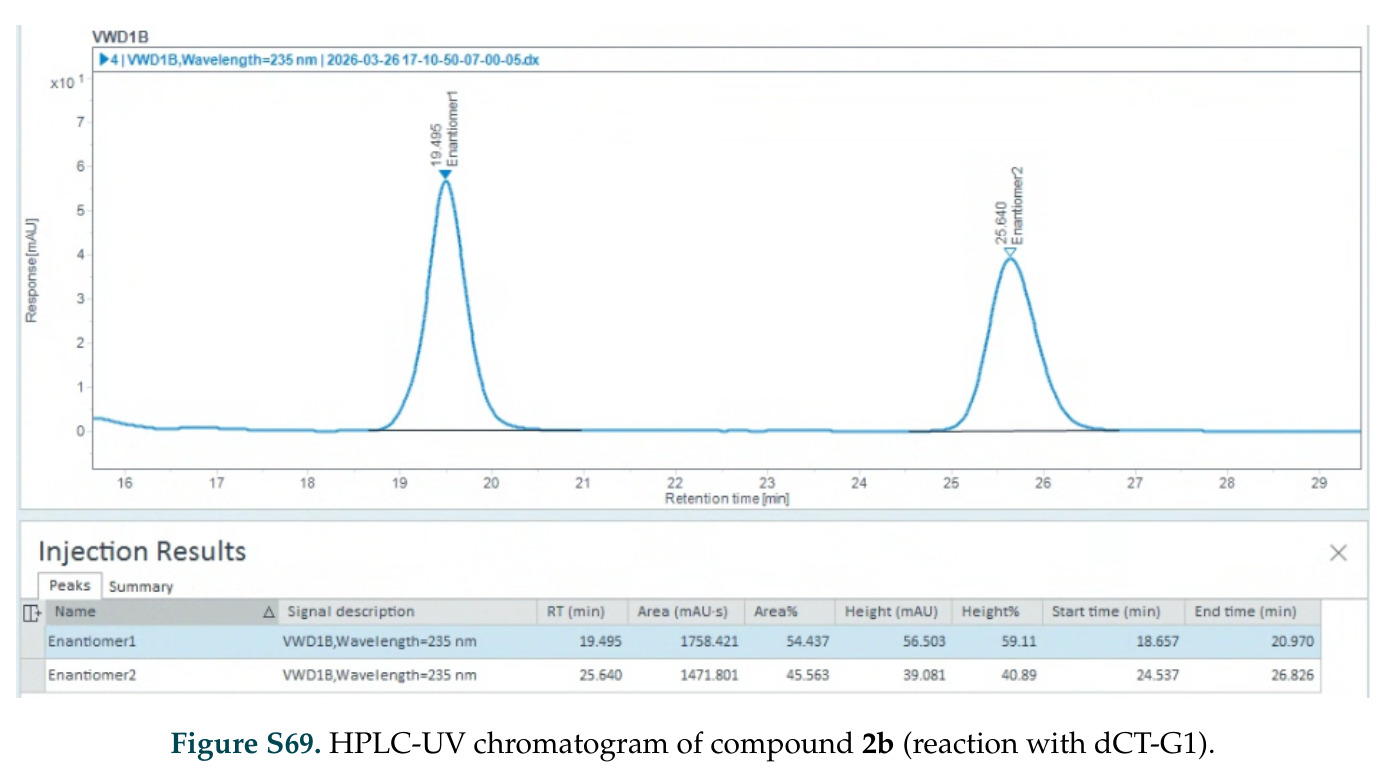
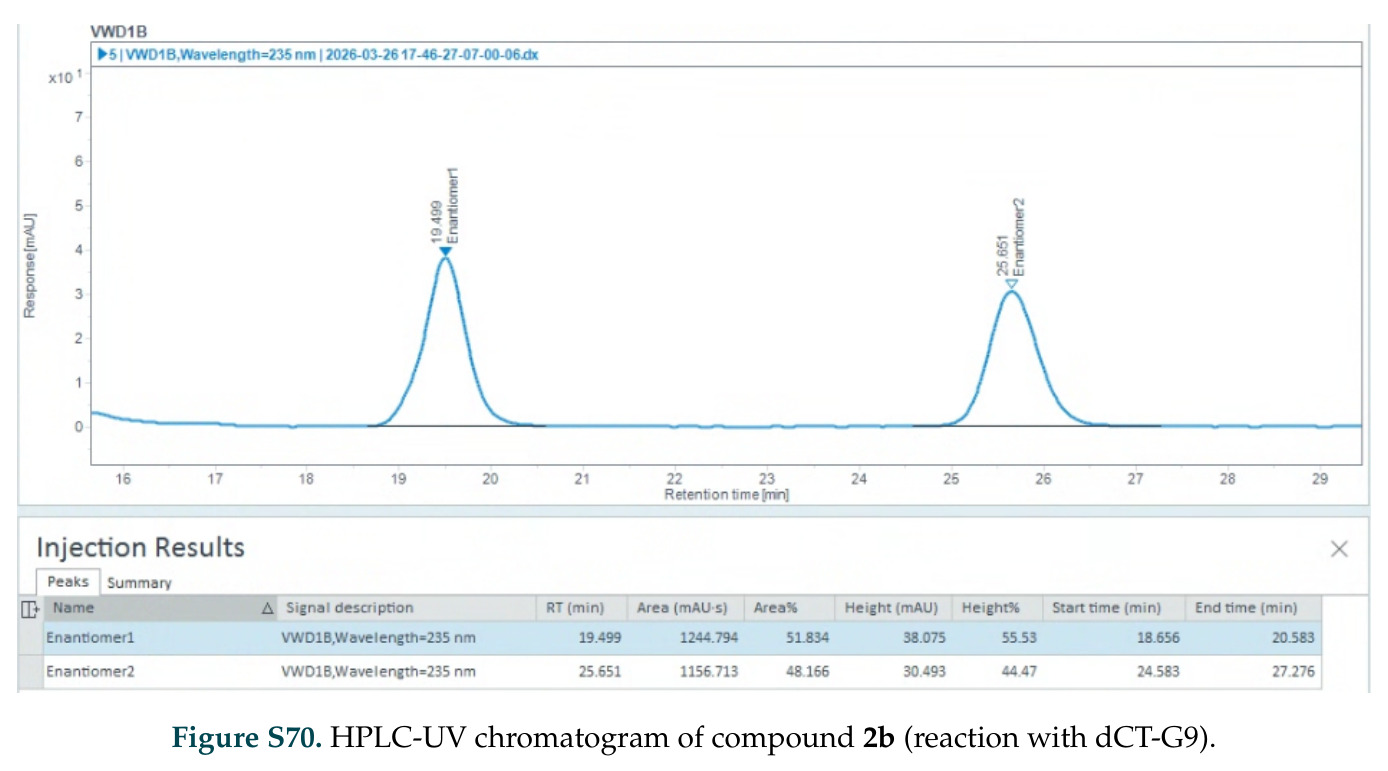
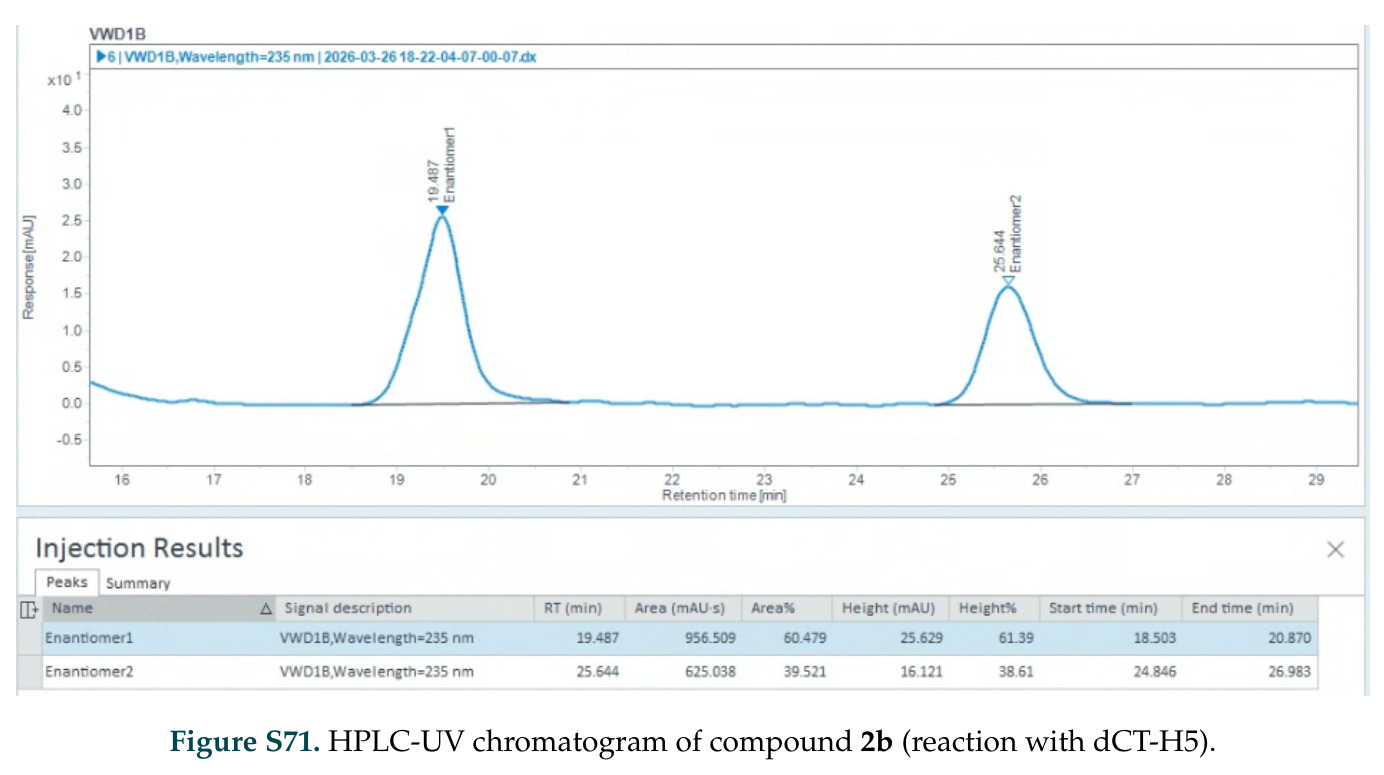
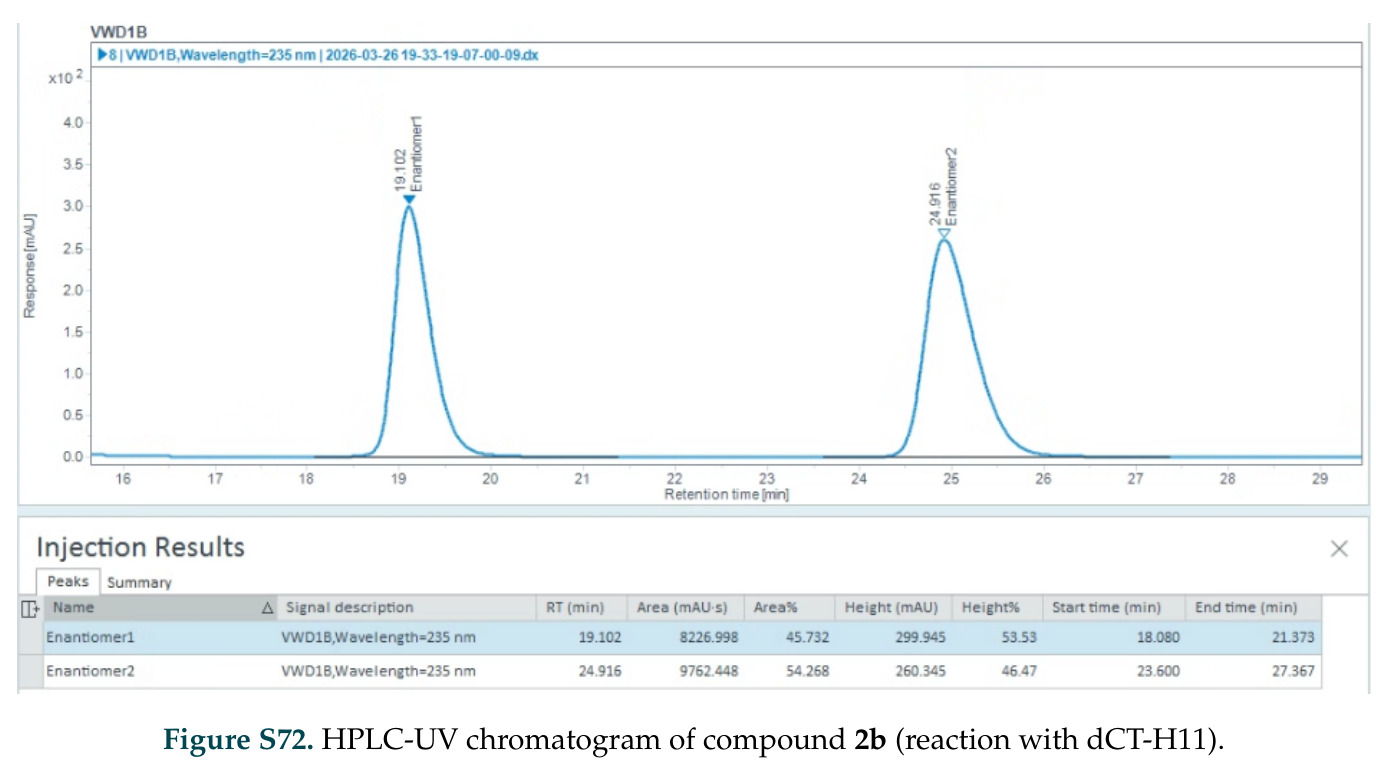
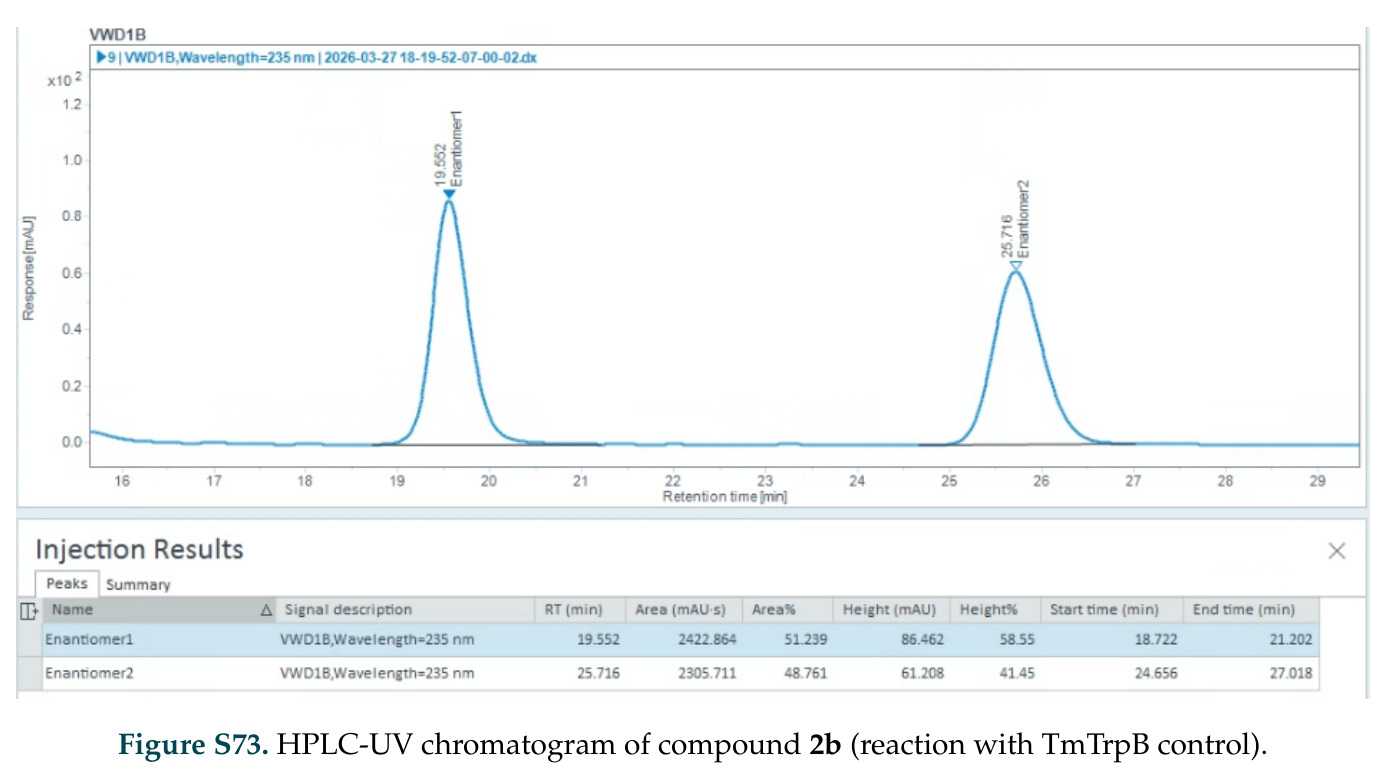
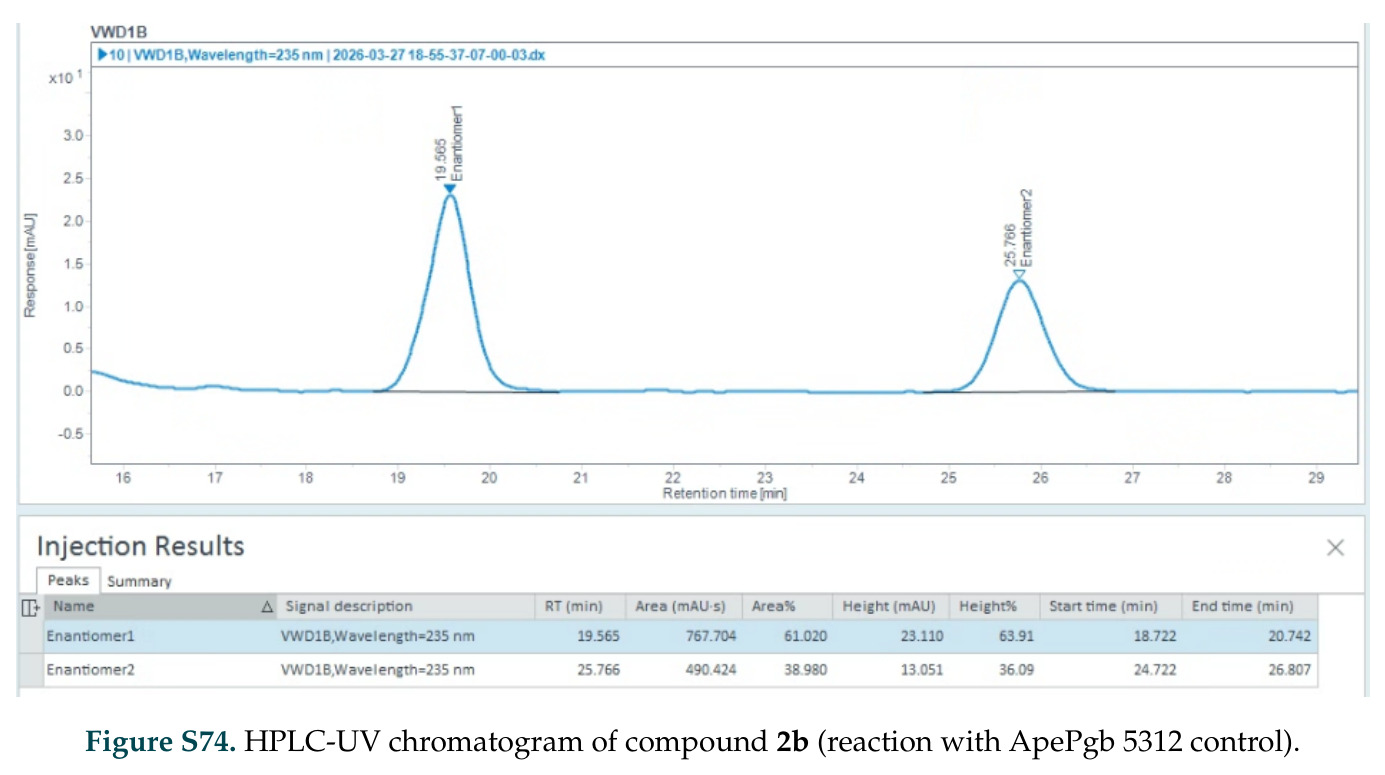
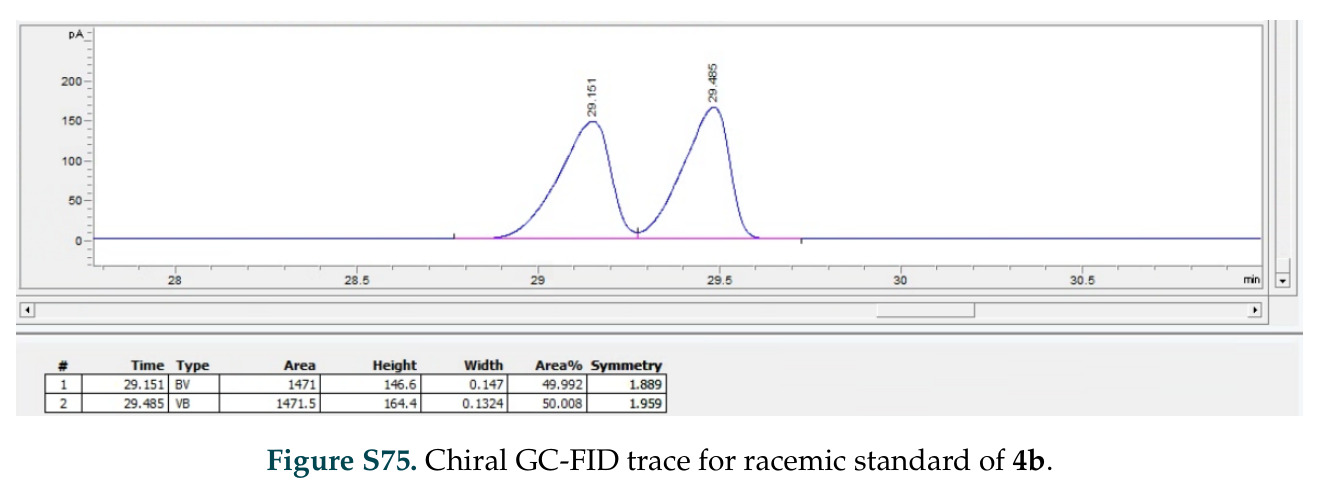
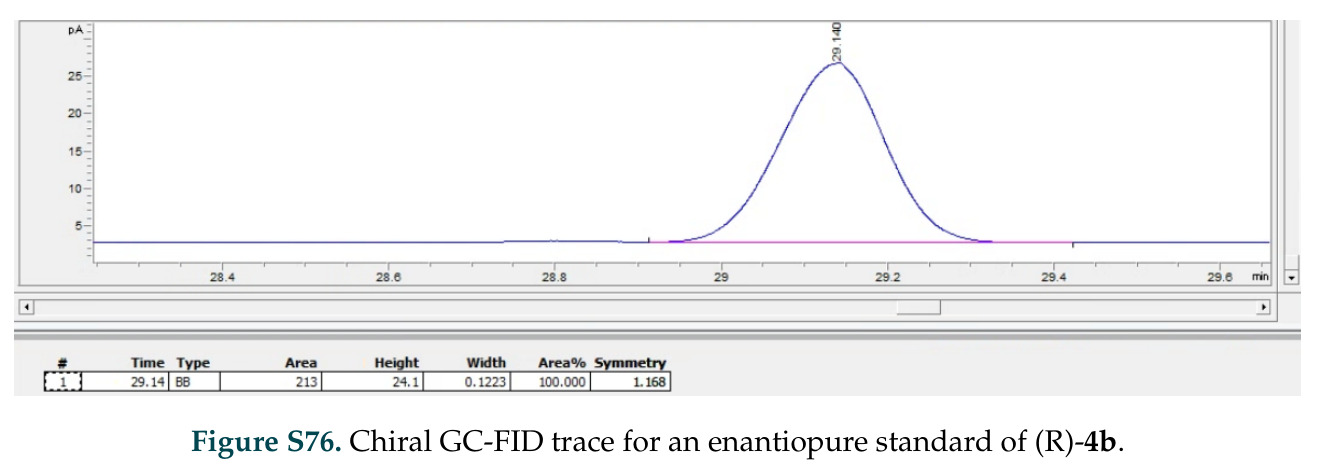
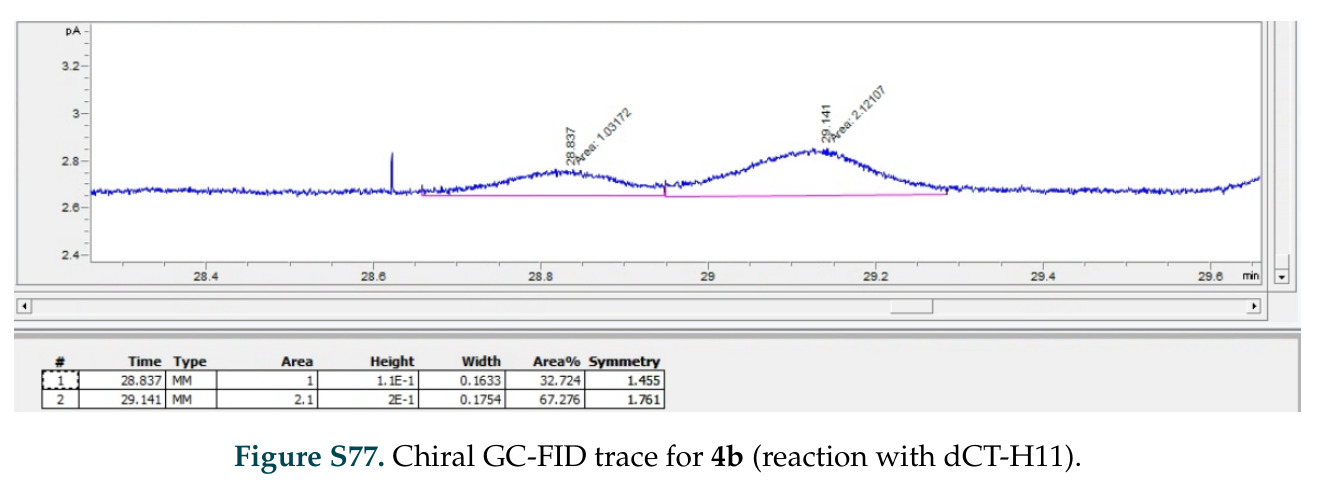
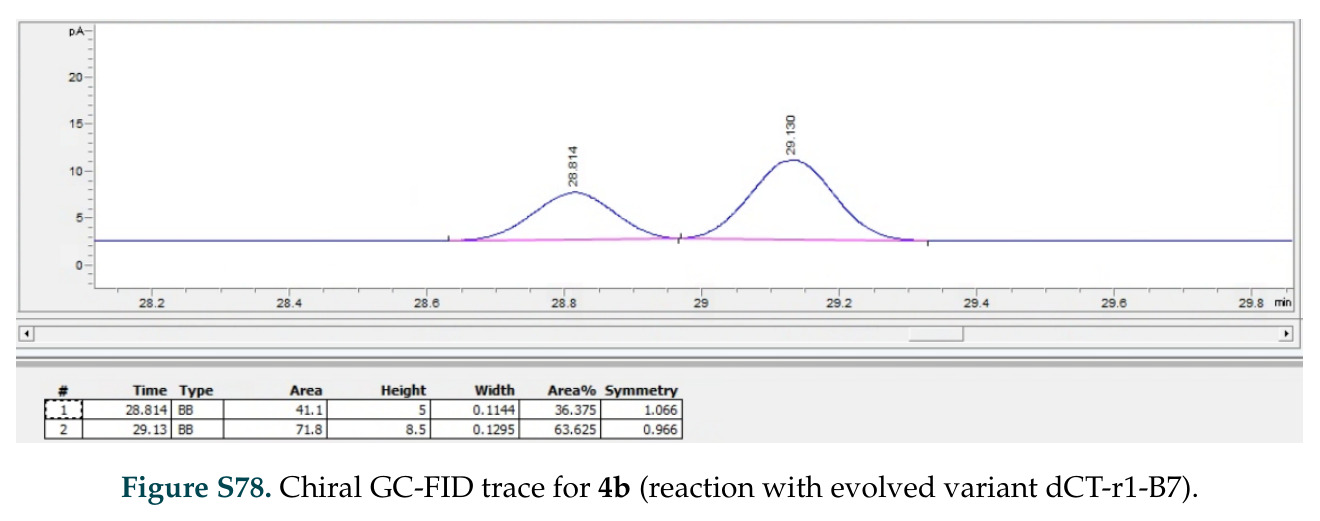
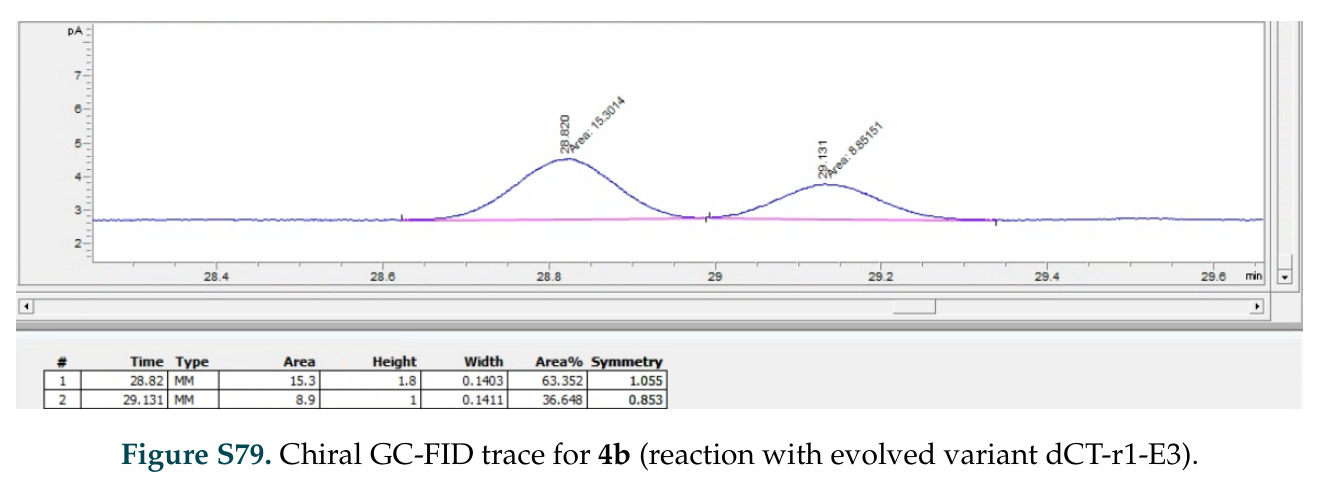
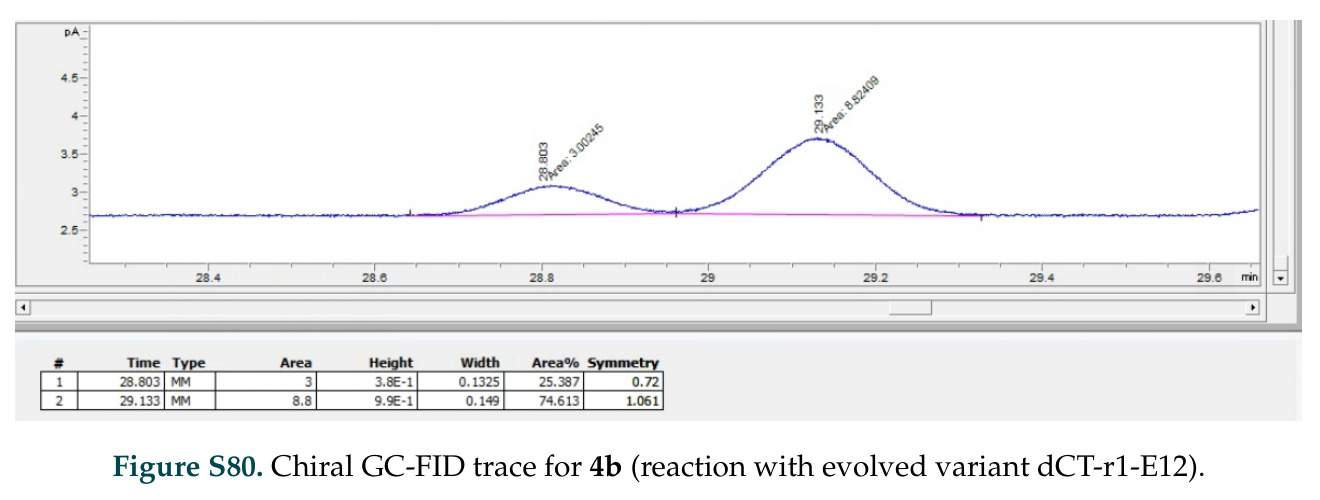
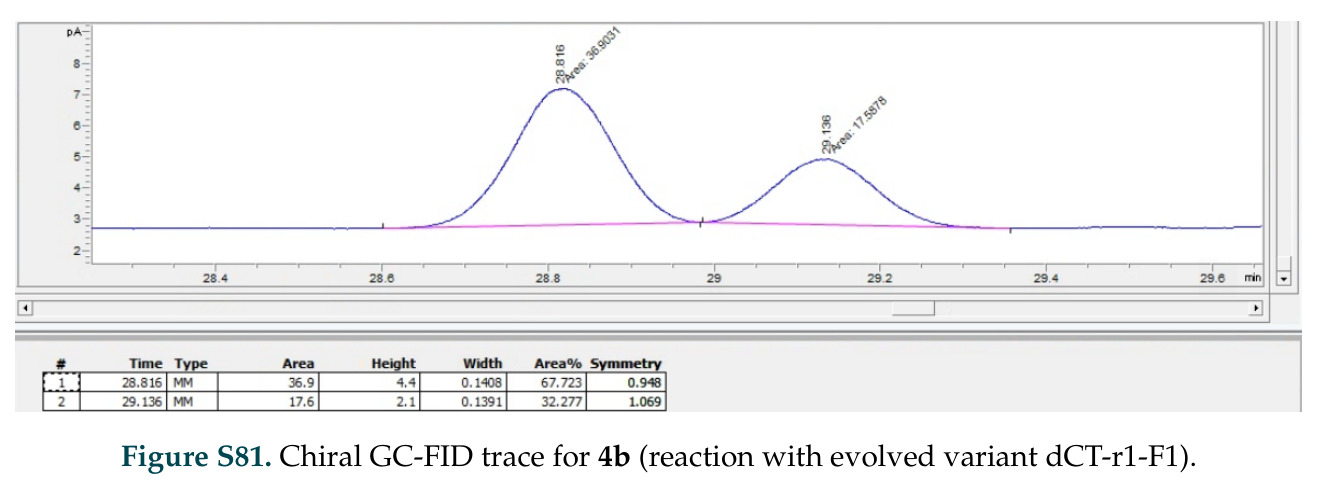
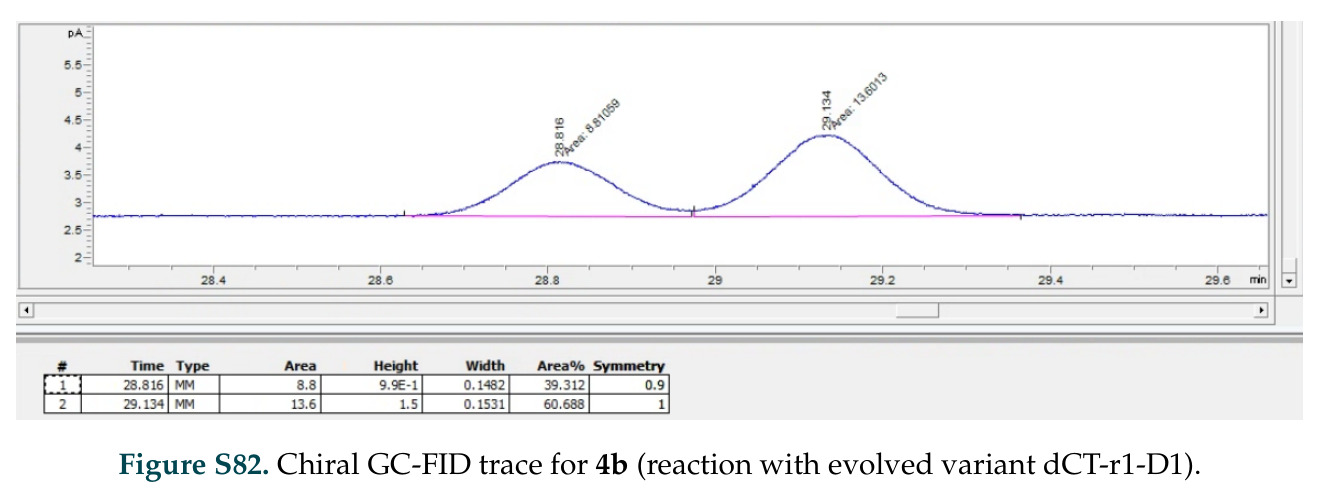
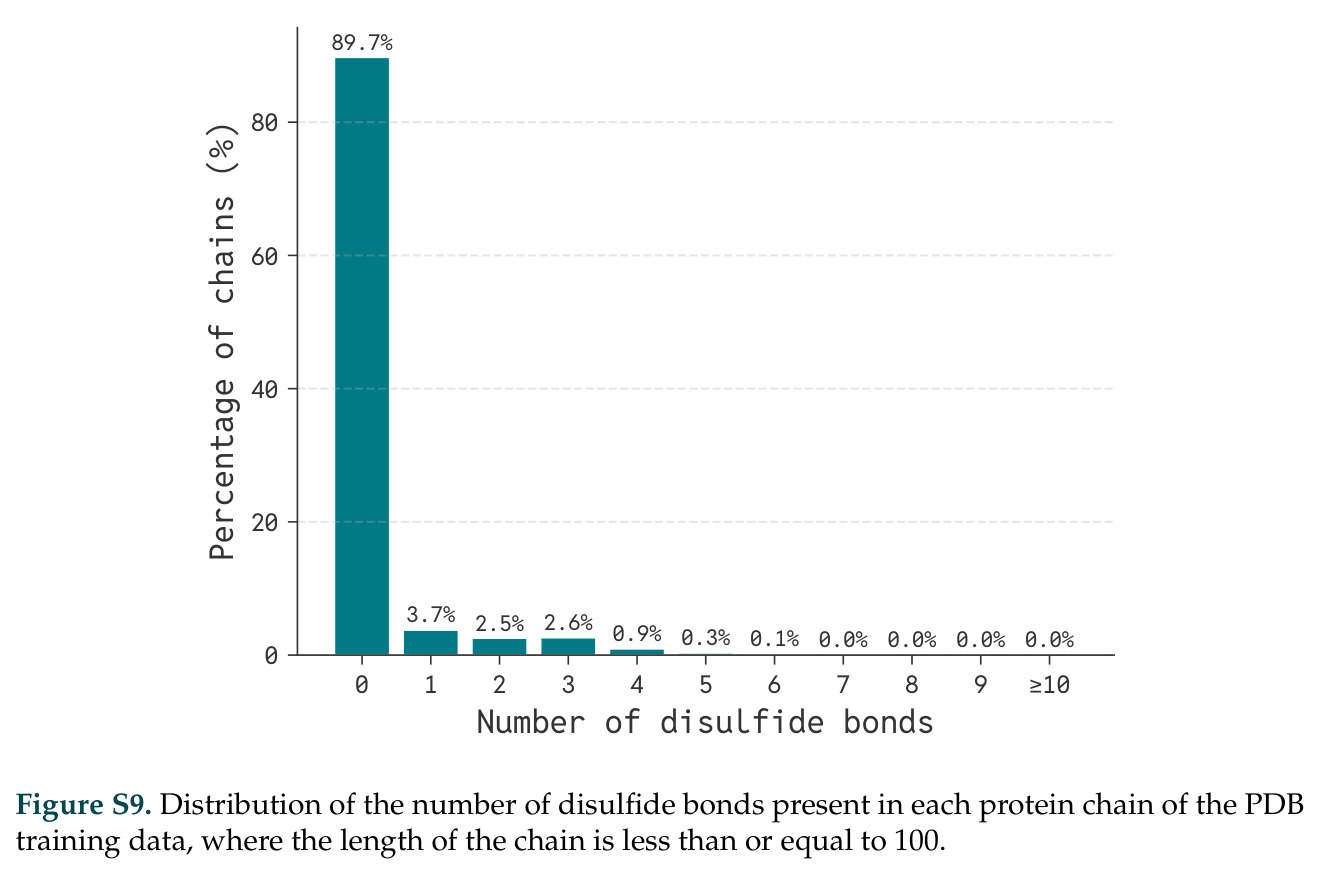
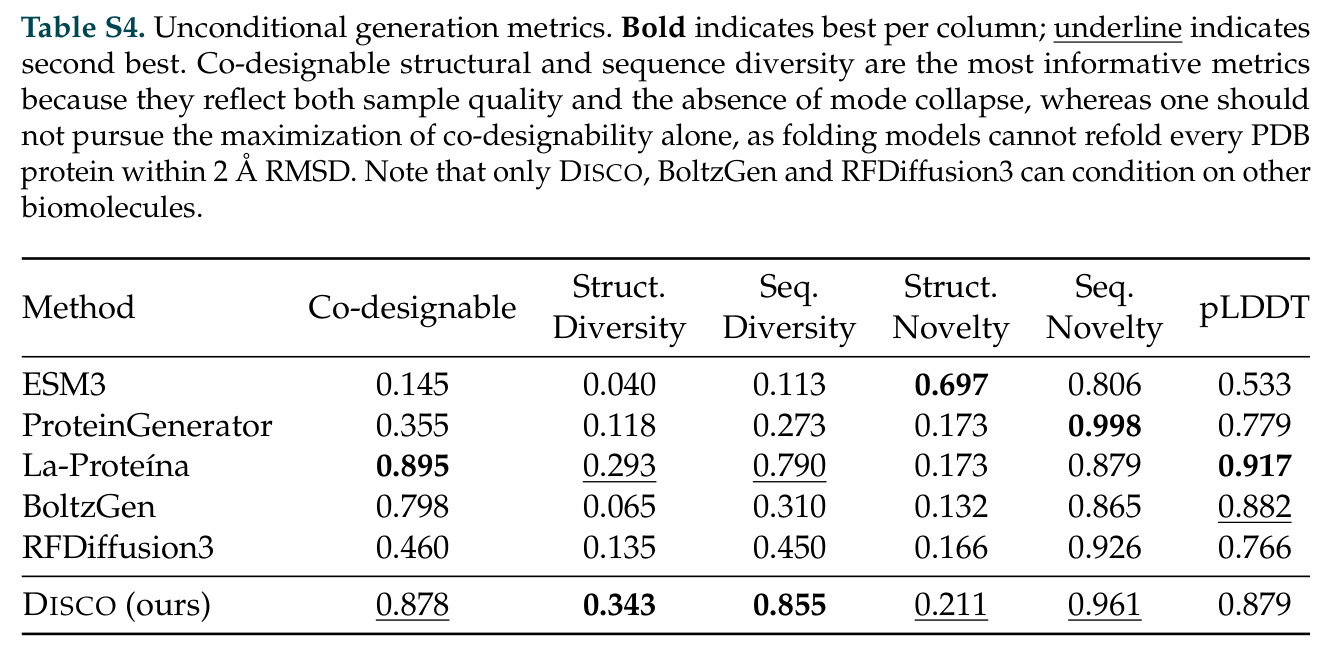
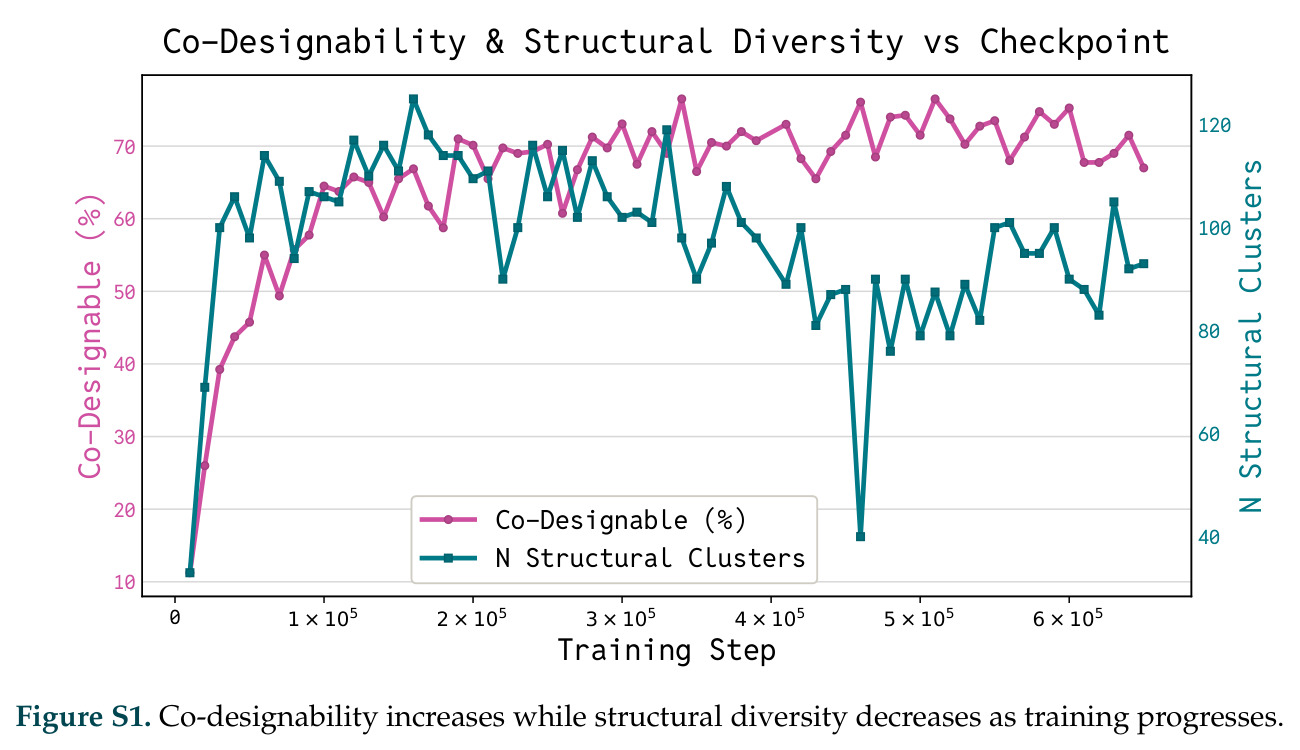
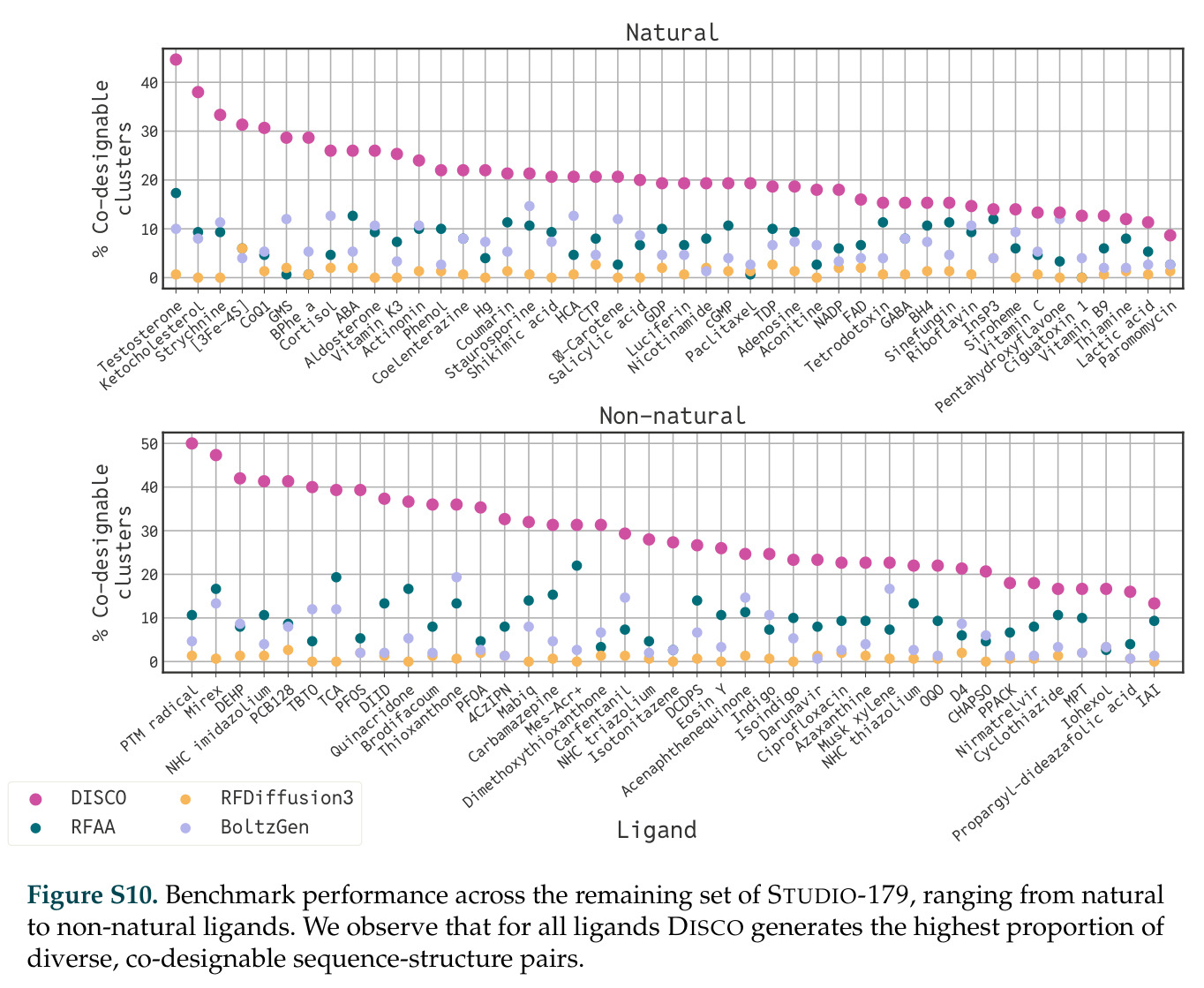
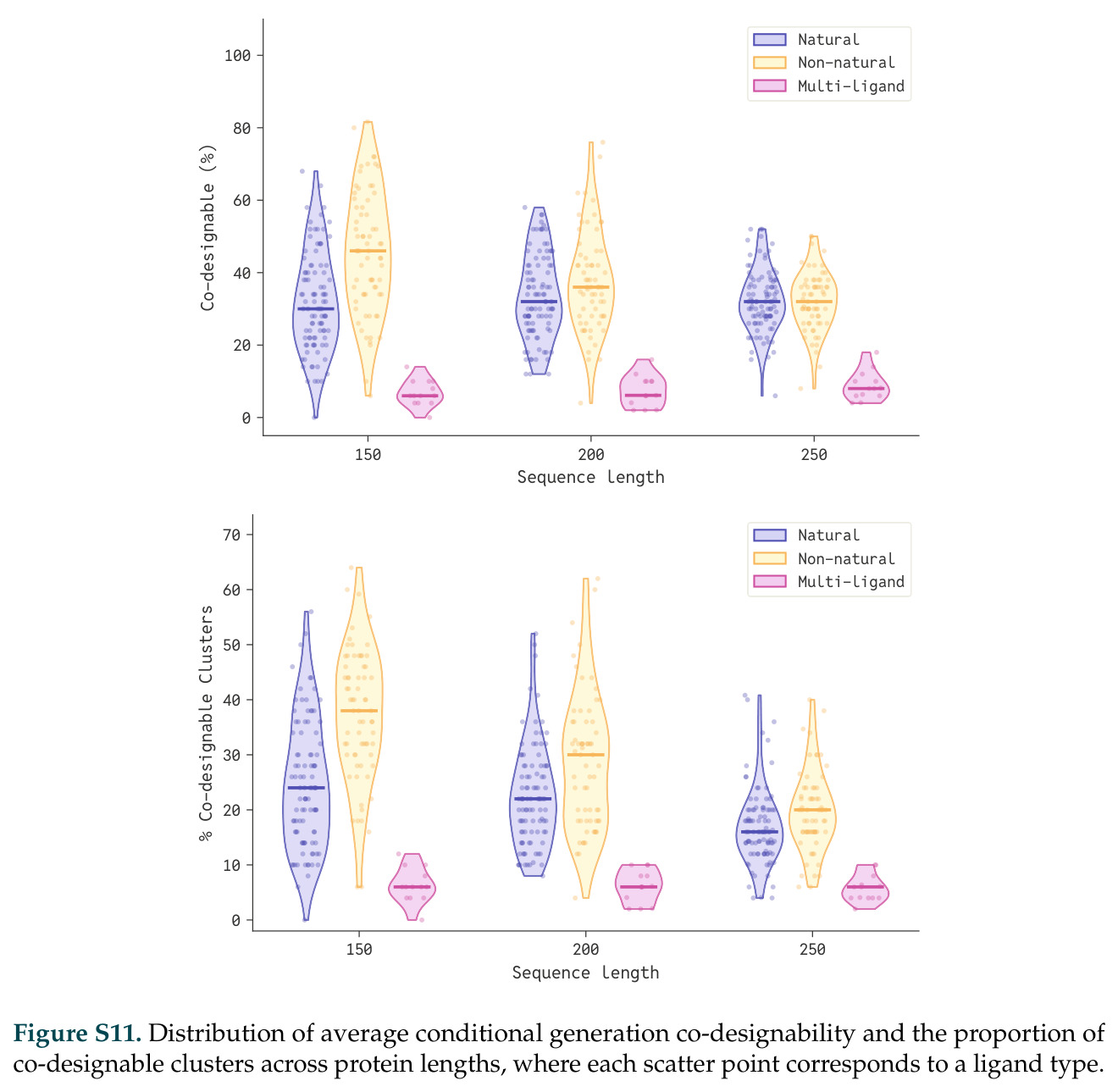
在计算评估方面,DISCO在无条件单体设计中表现优异:约90%的生成序列可以用ESMFold重折叠到设计骨架结构的2埃RMSD范围内,相比基线方法实现了最高的序列和结构多样性和新颖性,同时没有牺牲协同设计能力。在条件生成方面,DISCO在STUDIO-179基准测试中对179个配体中的178个生成了最高比例的多样化、可协同设计的复合物。推理时引导方面,FKC-MM生成的设计 enriched 了目标属性:在可设计的100氨基酸蛋白质中,top 2%包含6个二硫键,这一密度只有训练数据中top 0.2%的蛋白质才能匹配;FKC-SG在区分相似分子方面,生成了在目标和off-target复合物之间具有高分离度的蛋白质,优于best-of-N过滤方法。实验验证方面,从约一万到一万零四个计算生成的序列-结构对中,经过计算筛选得到90个设计进行湿实验测试。筛选结果显示设计的变体不仅具有功能(2.2-66%超过TmTrpB对照),而且在某些情况下表现出高催化活性:对甲氧基苯乙烯环丙烷化的最佳设计达到72%收率和4050总转化数,99比1非对映异构体比,超过早期进化的P411酶和近期设计的酶PNC2;B-H插入的最佳变体达到98%收率和5170总转化数,远超先前起点(120总转化数)和实验室进化变体(2490总转化数);C(sp3)-H烷基化的最佳设计达到42%收率和2360总转化数,与先前进化的P411-CHF催化剂(2030总转化数)相当;螺环丙烷化最具挑战性,只有适度活性。可进化性方面,对螺环丙烷化反应的dCT-H11进行一轮易错PCR和约700个突变体筛选,约35个变体显示出相对于亲本酶的明显改善活性,对映异构体选择性从35%对映体过量提高到49%对映体过量,甚至完全反转到负35%对映体过量,说明DISCO的设计占据具有可访问的上坡优化路径的序列空间区域。新颖性方面,最佳设计方案采用多种折叠和新颖 motif:dCT-H11的最接近结构匹配是来自极端嗜盐菌的TetR家族转录因子(PDB ID: 3CRJ,TM-score: 0.81),仅21%序列相似性;dCT-F9(最接近匹配: 5MSV,TM-score 0.52,5%序列相似性)和dCT-G9(最接近匹配: 7C7E,TM-score 0.51,9%序列相似性)采用与已知结构更远的折叠;在AlphaFoldDB中,基于五个最近活性位点残基,没有为其他设计识别到相应的 motif。所有最接近的结构匹配都不对应天然血红素结合蛋白。
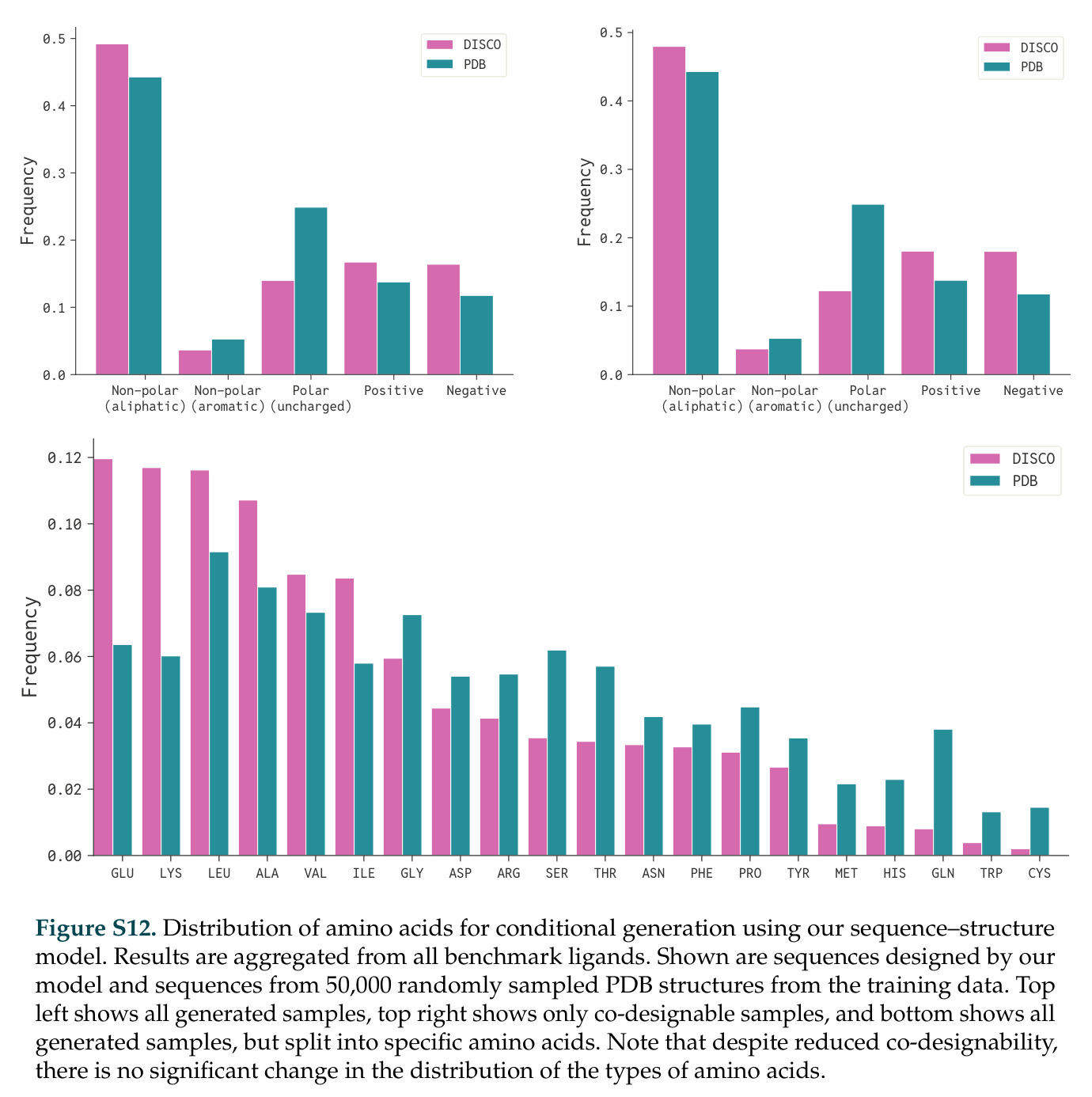
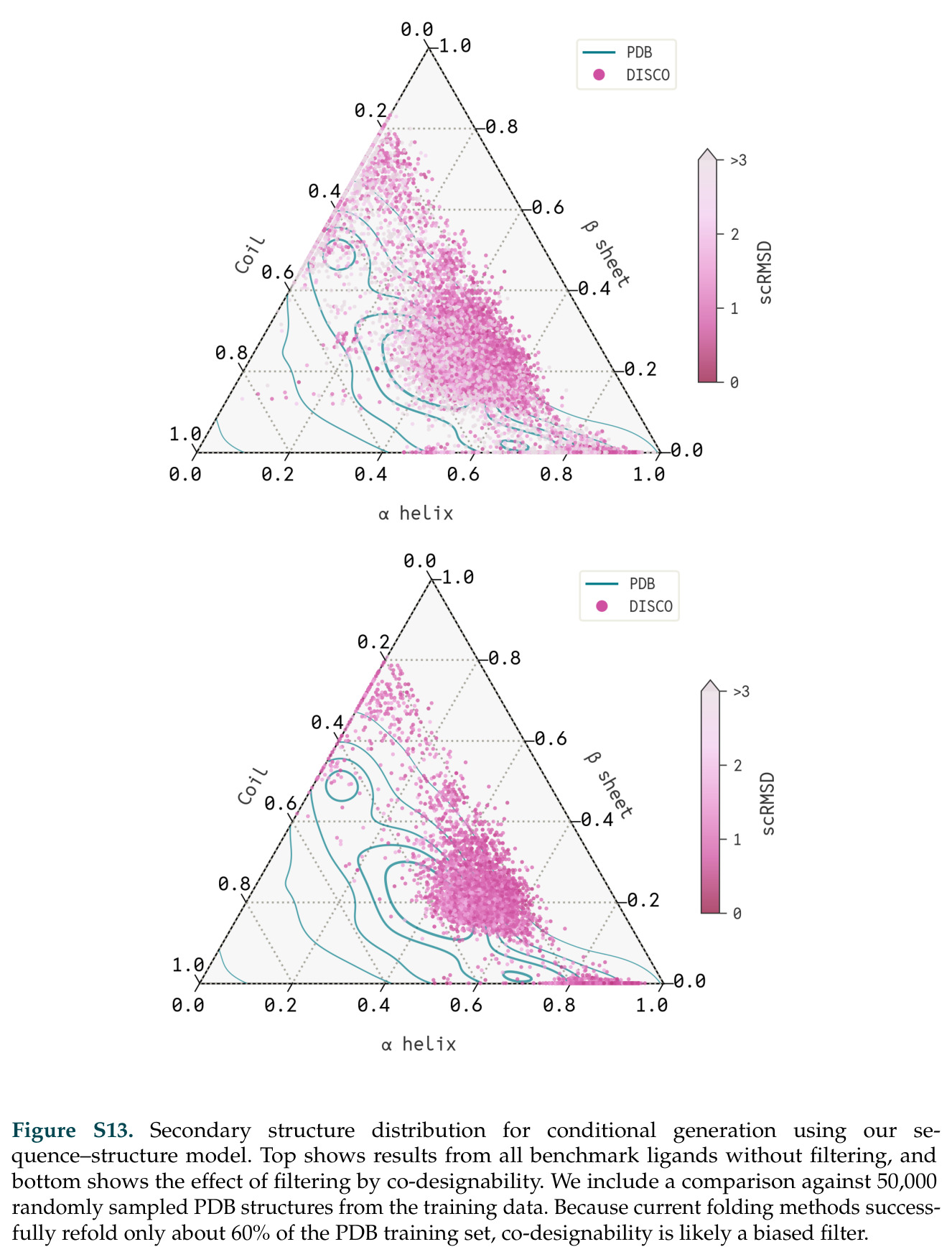
查看结构化数据
| 任务 | 指标 | 本文 | 基线 | 提升 |
|---|---|---|---|---|
| 无条件单体生成协同设计 | 重折叠到2埃RMSD内的比例 | 约90% | 未明确给出具体数字,但DISCO达到最高 | DISCO在序列和结构多样性和新颖性方面均达到最高,同时不牺牲协同设计能力 |
| STUDIO-179条件生成基准 | 可协同设计复合物的配体数量 | 179个中178个 | 未明确给出 | DISCO对几乎所有配体类型都能生成高质量复合物 |
| 对甲氧基苯乙烯环丙烷化 | 总转化数 | 4050 | Theozyme-based design PNC2: 630, Evolved P450H2-5-F10: 364 | 超过PNC2 6.4倍,超过进化酶11倍 |
| B-H插入 | 总转化数 | 5170 | Evolved Rma cyt c: 2490, Rma cyt c: 120 | 超过进化变体2.1倍,超过起点43倍 |
| C(sp3)-H烷基化 | 总转化数 | 2360 | Evolved P411-CHF: 2030 | 与进化变体相当,而无需14轮定向进化 |
| FKC-SG特异性引导 | 目标和off-target复合物分离度 | 高分离度 | best-of-N过滤在某些情况下不产生通过计算机过滤的命中 | FKC-SG在best-of-N失败的情况下仍能生成特异性结合蛋白质 |
局限与改进
作者承认的局限性包括:当前评估由于DISCO中冻结的pLM没有在多聚体复合物上训练,被限制在非蛋白质结合任务;螺环丙烷化反应表现出更高的背景活性和更低的可设计活性,这表明某些类型的转化仍然具有挑战性;方法依赖于奖励函数的质量,需要仔细的标准化函数来平衡序列和结构贡献;训练和推理需要大量计算资源,尤其是对于复杂的多模态生成。我自己观察到的局限性:虽然DISCO生成的酶显示出高活性,但大多数实验验证的反应仍然有限(四种底物),更广泛的反应类型和更复杂的化学转化尚未验证;虽然DISCO可以避免预定义催化残基,但它仍然依赖于反应中间体的DFT计算几何,这可能不适用于机制完全未知的反应;研究没有提供系统分析来说明DISCO的设计与天然血红素蛋白相比,在进化稳定性和表达水平方面的差异;FKC-SG在极端相似分子如对映异构体上的表现尚未评估,这可能对药物设计等应用很重要。
独立分析的弱点
DISCO的一个独立分析弱点是其计算资源需求较高,训练单一深度神经网络处理多模态扩散需要大量GPU内存和计算时间,这可能限制其在资源受限实验室的广泛应用。改进方向包括模型蒸馏、轻量化架构如将Pairformer堆栈替换为更高效的结构、以及量化技术。另一个弱点是在处理大分子复合物时的性能限制:由于pLM没有在多聚体上训练,DISCO当前无法可靠地生成蛋白质-蛋白质界面。改进方向包括使用专门的多聚体语言模型或微调现有pLM在多聚体数据上。第三个弱点是FKC框架依赖于可微的奖励函数,而许多重要的蛋白质性质如稳定性、表达水平难以精确建模。改进方向包括使用代理模型或贝叶斯优化来整合这些难以建模的目标。第四个弱点是当前DISCO主要关注静态结构设计,没有明确考虑动力学特性如柔性、构象变化。改进方向包括整合分子动力学模拟或使用变分扩散过程来建模构象分布。
未来方向
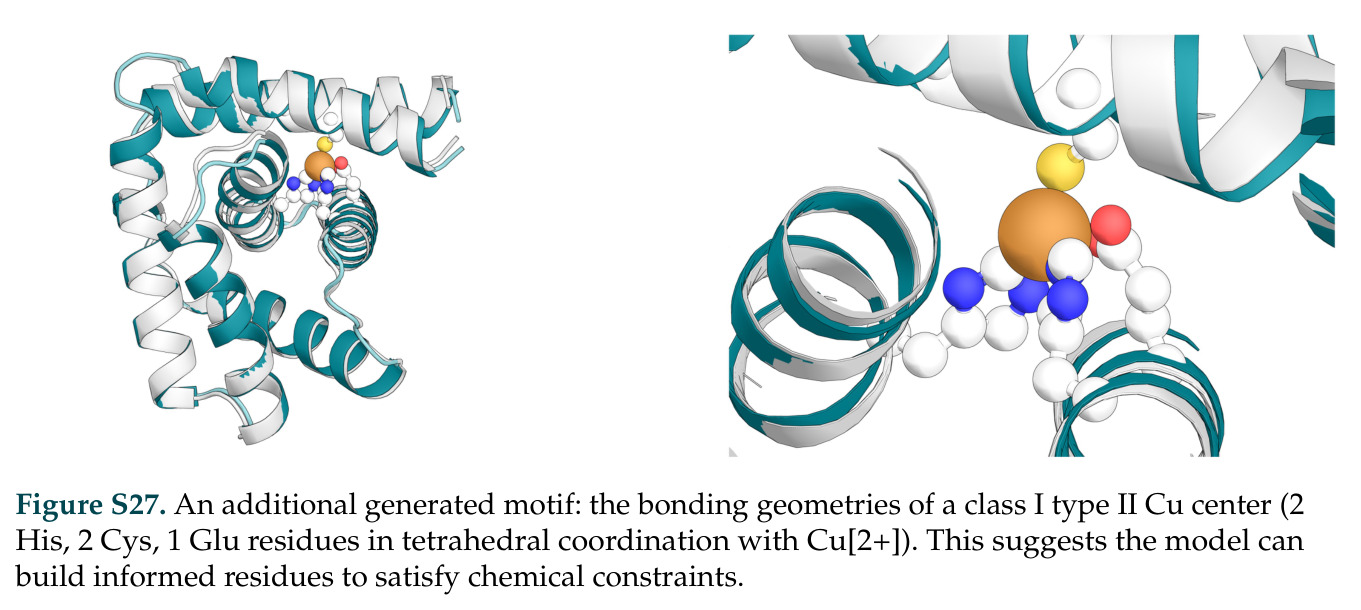
作者提出的未来工作方向包括:掌握日益复杂的机制可能需要关闭生成发现和定向进化之间的循环,并通过更丰富的生物物理约束进行通知。基于DISCO的成果可以延伸的方向包括:扩展到更多类型的反应,当前工作聚焦于卡宾转移反应,但DISCO的通用框架应该适用于其他新化学反应类型,如碳氮键形成、氧化还原反应等;整合更精确的生化约束,包括静电互补、疏水匹配、氢键网络等更细致的物理化学约束,以提高设计的精确性和成功率;迭代优化循环,将DISCO的生成能力与定向进化结合,形成一个反馈循环,其中进化数据用于重新训练或微调模型,从而逐步改进设计质量;多目标优化,扩展FKC框架以同时优化多个目标,如活性、选择性、稳定性、表达水平等,这对于实际应用至关重要;扩展到蛋白质-蛋白质界面设计,通过在多聚体数据上训练pLM或引入专门的界面建模模块,使DISCO能够生成功能性蛋白质复合物;整合实验筛选数据,将高通量筛选结果反馈到模型中,通过主动学习或贝叶斯优化来指导生成过程,减少搜索空间;设计新型催化机制,不仅限于血红素酶,还可以探索其他辅因子如铜、锌、钴或全新的催化机制,扩展DNA编码化学转化的范围。
复现评估
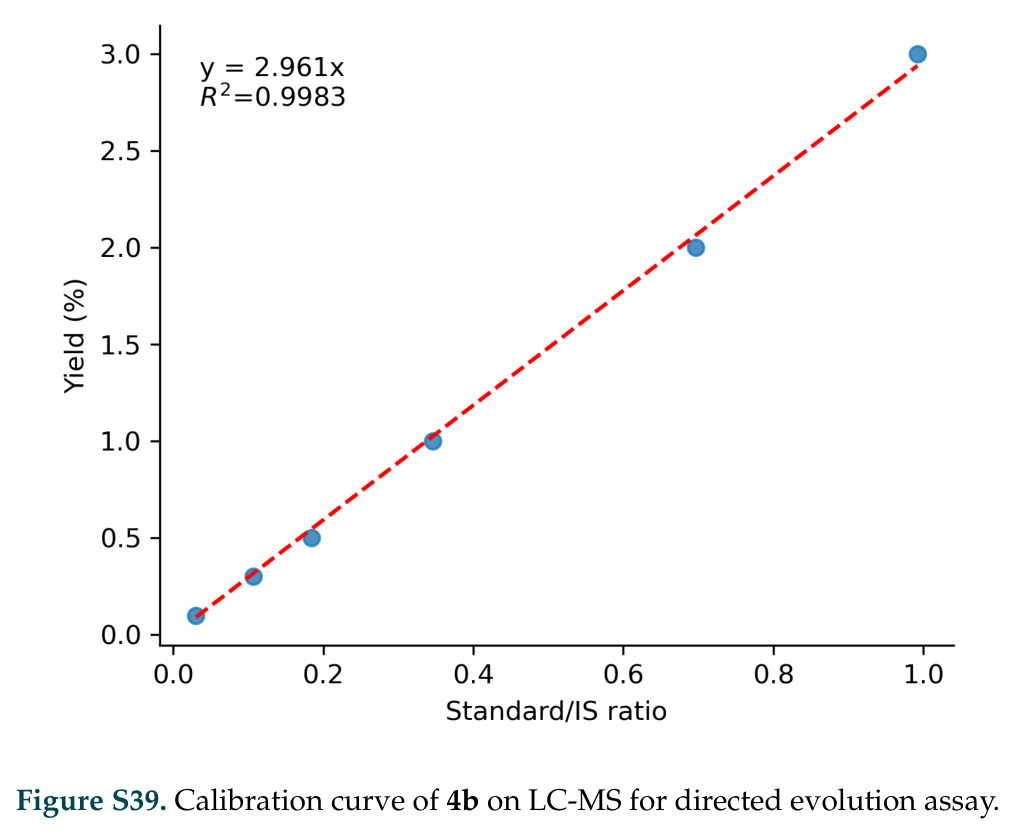
复现评估方面,DISCO的代码和模型已在GitHub开源,这大大降低了复现门槛。研究使用了公开的Protein Data Bank数据,无需特殊过滤,避免了选择偏差。实验验证的90个基因序列在论文补充材料中提供,包括具体的筛选方案和反应条件。计算细节如模型架构、训练超参数、推理策略在论文和补充材料中有详细描述,这有助于复现计算结果。然而,复现仍面临一些挑战:训练完整的DISCO模型需要大量GPU资源(虽然具体资源需求未明确说明,但考虑到多模态扩散的复杂性,可能需要多张高端GPU);湿实验验证需要分子生物学设备和专业知识,包括基因合成、蛋白质表达、活性筛选等;实验验证使用了多种分析技术如高效液相色谱、核磁共振、气相色谱质谱,这些设备和专业知识可能不是所有实验室都具备;反应中间体的DFT计算需要量子化学软件如Gaussian或ORCA和相关专业知识。总体而言,DISCO的复现性处于较高水平,特别是计算部分可以通过开源代码和数据复现,但完全复现包括实验验证需要相应的资源。
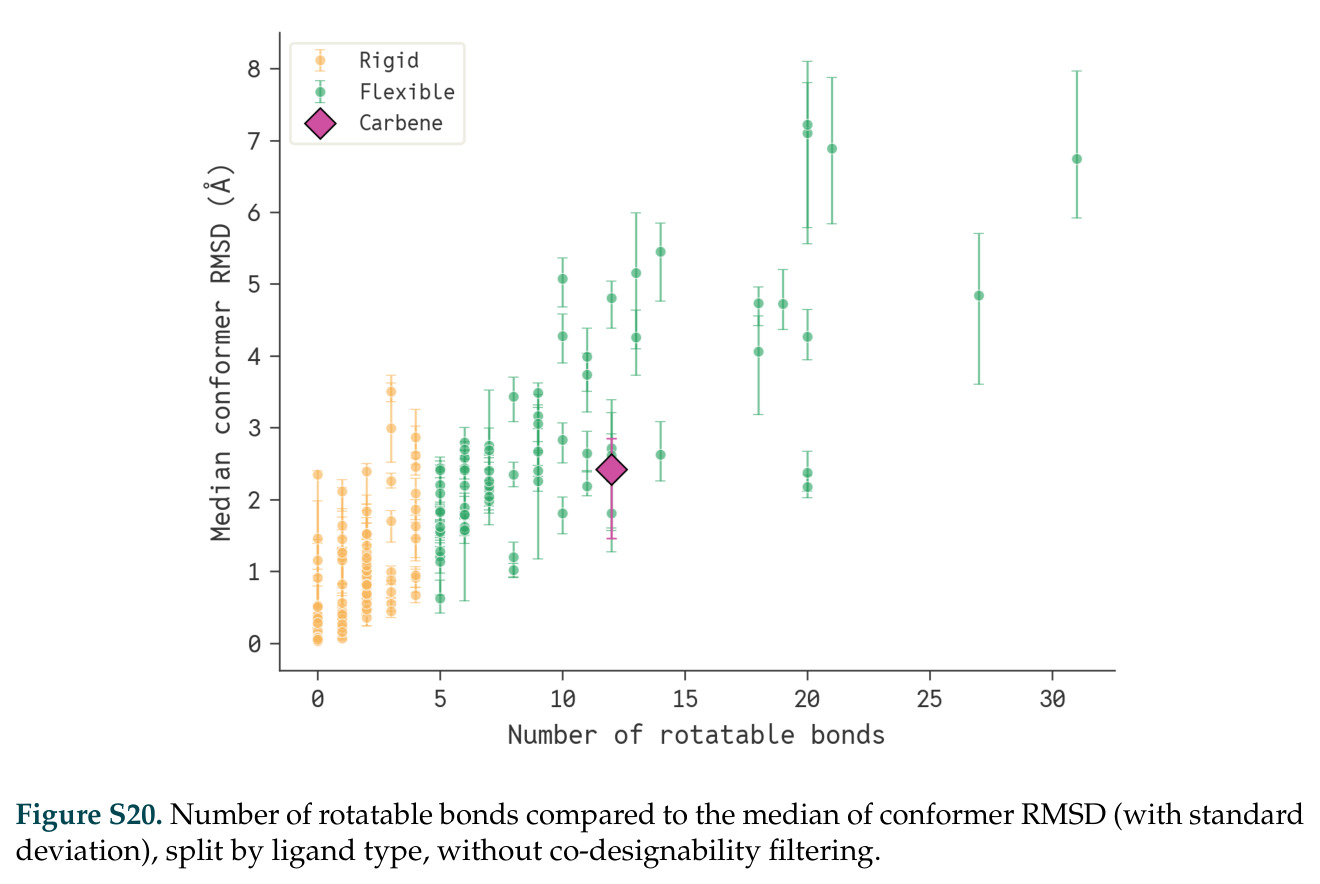
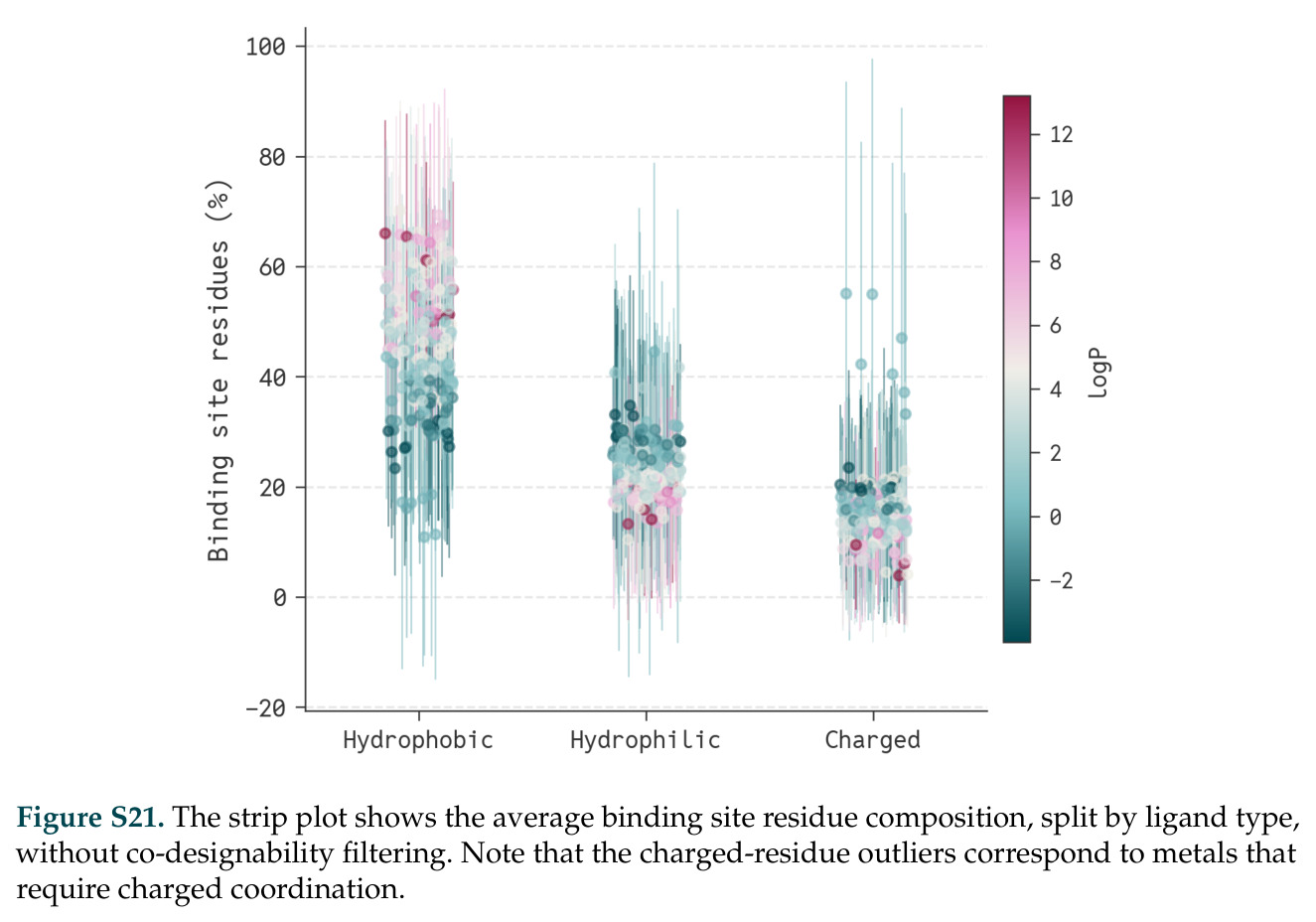
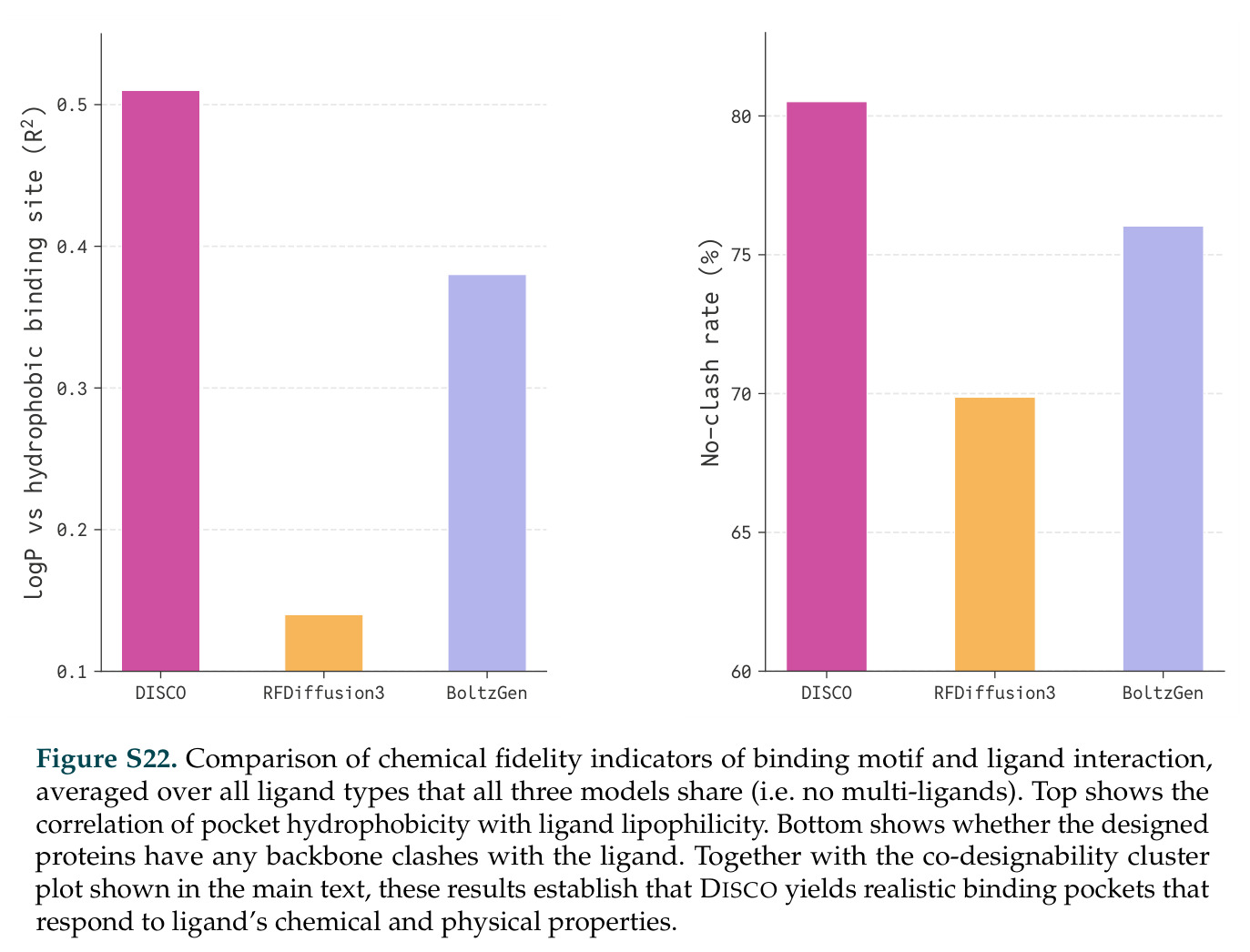
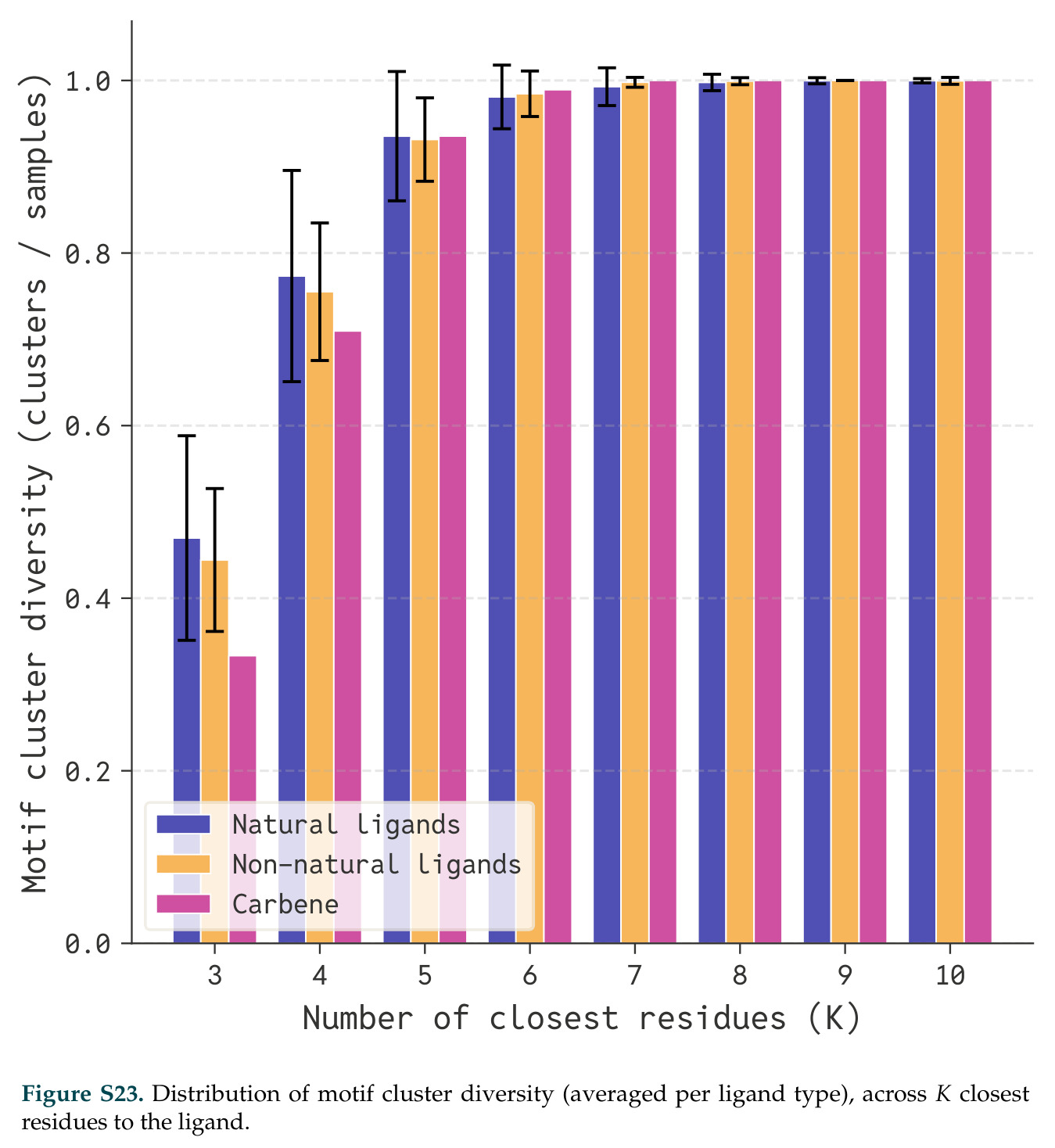
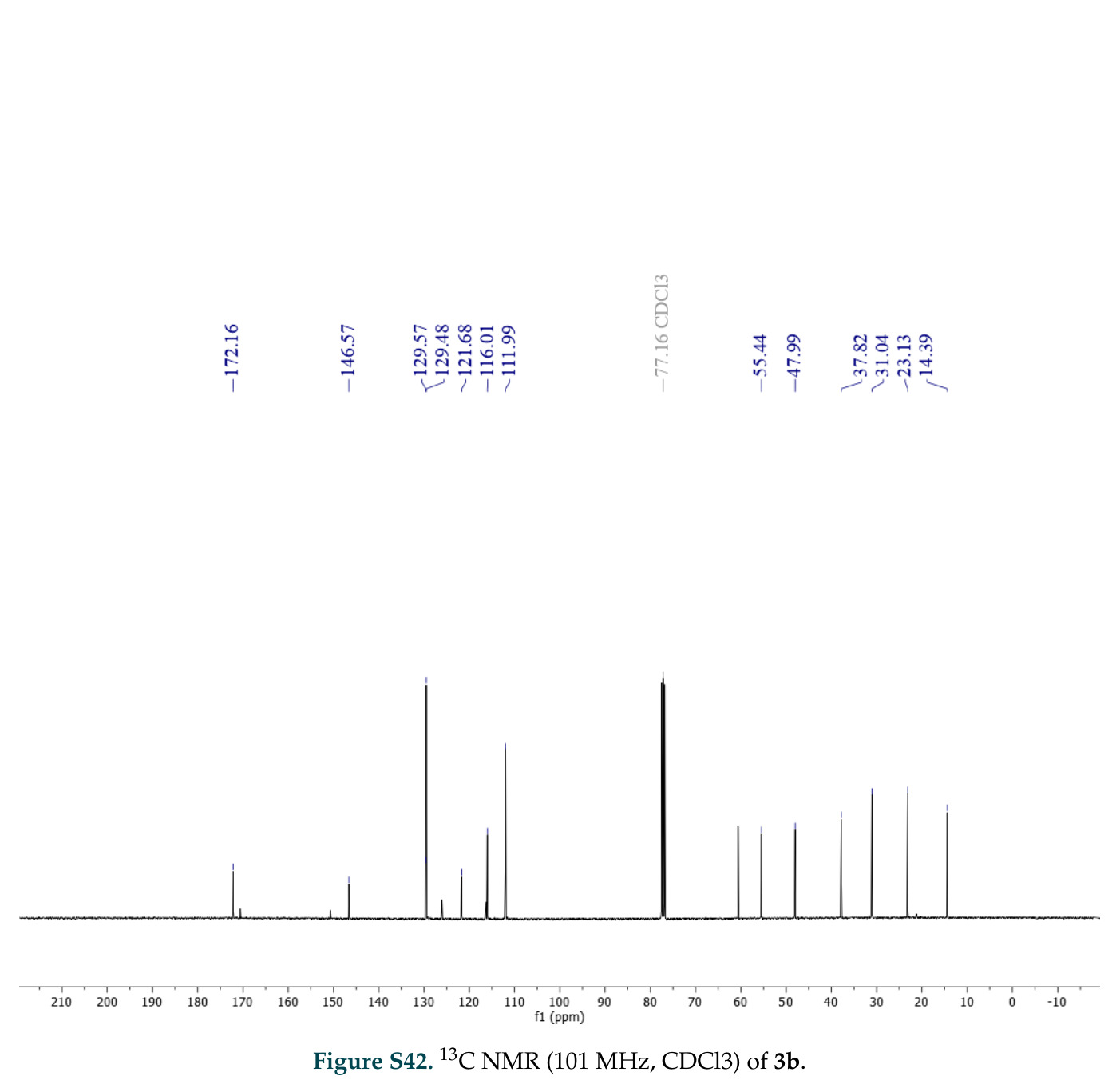
论文图表