面向生物图像的自动图像级形态特征标注 Automatic Image-Level Morphological Trait Annotation for Organismal Images
用稀疏自编码器定位昆虫部位,再让多模态大模型生成可解释的形态特征描述。
前置知识
稀疏自编码器 (Sparse Autoencoder, SAE)
SAE 是一种把高维密集表示解耦成稀疏、人类可解释特征的字典学习方法。它通过编码器把输入向量 $z \in \mathbb{R}^d$ 映射到高维潜空间 $u \in \mathbb{R}^n$,用 ReLU 强制稀疏性,再用解码器线性重建 $z$。训练目标是 $\mathcal{L} = \|z - \tilde{z}\|_2^2 + \alpha R(g(z))$,其中 $\alpha$ 控制稀疏惩罚。重要的是,'稀疏 + 非负'双重约束使得每个 latent unit 倾向于只对单一语义概念响应(monosemantic),从而成为天然的'部件检测器'。
本文的核心创新就是把 SAE 用作无监督的部位提议器,是把可解释性研究从语言模型迁移到生物视觉的关键桥梁。
形态特征 (Morphological Trait)
形态特征是生物体可测量的物理属性,如翅长、腿节比例、复眼间距、体表纹理等。在生态学中,特征比物种名更能预测物种对环境变化的响应——研究表明形态特征对生态位的预测准确率可达 85%。但人工测量每件标本耗时数分钟,全球 30 亿+ 件自然史标本的完整特征普查需要消耗'人-世纪'级别的工作量。
这是本文要解决的生态学痛点,特征是连接'物种鉴定'和'功能生态学'的机制纽带。
DINOv2 与自监督视觉骨干
DINOv2 是 Meta 训练的自监督 ViT 模型,其 patch-level 特征对语义部位具有较强的空间一致性。本文用其第 10 层输出作为 SAE 输入,并在 1000 物种基准上对比了 CLIP/SigCLIP/DINOv2 的 kNN 分类准确率:DINOv2 达到 41.28%,远超 CLIP ViT-B/16 的 24.57%。
选 DINOv2 不是随意的,而是因为其特征对生物细粒度部位的捕捉能力远超 CLIP 类的对比学习骨干。
多模态大语言模型 (MLLM)
MLLM 能同时理解图像和文本并生成自然语言描述。本文主要使用 Qwen2.5-VL-72B(开源,可本地部署)和 GPT-5-mini(闭源 API)作为'特征语言化器'。关键实验发现:72B 模型比 7B 模型少幻觉(更少把背景错误识别为身体部位),GPT-5-mini 的人工评分最高(4.04 分)。
MLLM 不是简单的'描述生成器',而是整个流水线最后一道质量关——它的规模直接决定特征描述是否可信。
BIOSCAN-5M 数据集
BIOSCAN-5M 是 2024 年 NeurIPS 发布的多模态昆虫标本数据集,包含图像、DNA 条形码、分类、地理和体型信息。其中仅 9.2% 的样本有物种级标注,但这种'物种级监督'正是本文稀疏性筛选和 species-contrastive ranking 所需的关键信号。
理解数据规模(百万级图像但只有 9.2% 物种标注)才能体会本文'用稀疏自编码器压缩无标注特征 + 用物种标签过滤'的设计合理性。
研究动机
生物多样性危机下,生态学家迫切需要大规模形态特征数据来理解物种对环境变化的响应,但全球 30 亿+ 件自然史标本的完整特征普查需要消耗'人-世纪'级别的人工劳动——即使测量最简单的体长或胫节比,每件标本也要数分钟。现有自动化方案同样捉襟见肘:Inselect 这类工具最终把 GUI 扔回给用户,让人工每张图像还要花 108 秒纠正边界框;针对腊叶标本的 50+ 篇论文综述指出,看似简单的'叶面积''叶缘类型'任务在每个数据集上都需要重新做数据增强和调参,没有任何方法能干净地跨馆迁移。生物学'跨分类群异质性'(从被子植物叶片到黄蜂触角,特征流形会剧变)、不可控的姿态、标本保存伪影、背景杂乱这四重难题叠加,使标准监督学习在标签稀缺、形态非平稳、目标仅占画面微小区域的最坏情况下完全失效。
本文的目标是本文提出一个端到端、可扩展的形态特征自动标注流水线,能在没有人工逐件测量的前提下,把无标注的标本图像转成'图像-特征描述'对,并在 BIOSCAN-5M 上构造出覆盖 736 物种、417 属、19K 张昆虫图像、平均每张 4.2 条、共 80K 条特征描述的 BIOSCAN-TRAITS 数据集。最终用这些机器生成的特征去微调 BioCLIP,使零样本物种分类在 in-the-wild 的 Insects 基准上获得显著提升。
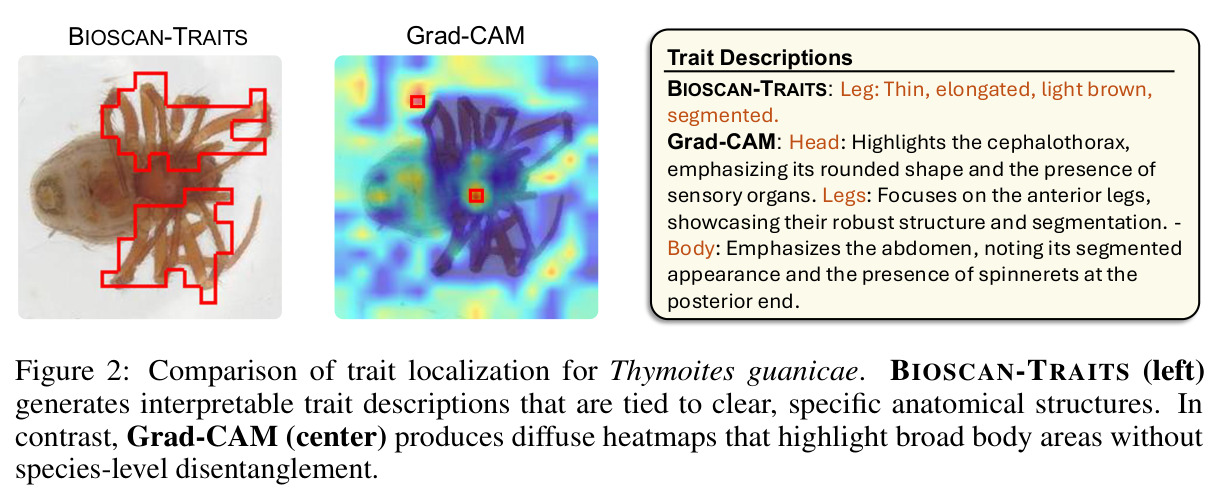
与已有工作不同的是,已有自动化尝试走两条路:要么用 Grad-CAM 类显著性方法给出弥散热力图,让 MLLM 描述整张图——但 MLLM 会陷入'描述整个场景'的幻觉;要么用有监督部件检测器——但需要昂贵的人工部件标注。本文的关键洞察是'可解释性研究里的稀疏自编码器可以被借用作无监督部位检测器',配合物种对比排序(focal 物种激活强但同属近缘物种弱)天然解决了'跨分类群异质性 + 无需人工部件标签'的难题,这是之前任何自动化方案都没有利用过的切入口。
核心方法
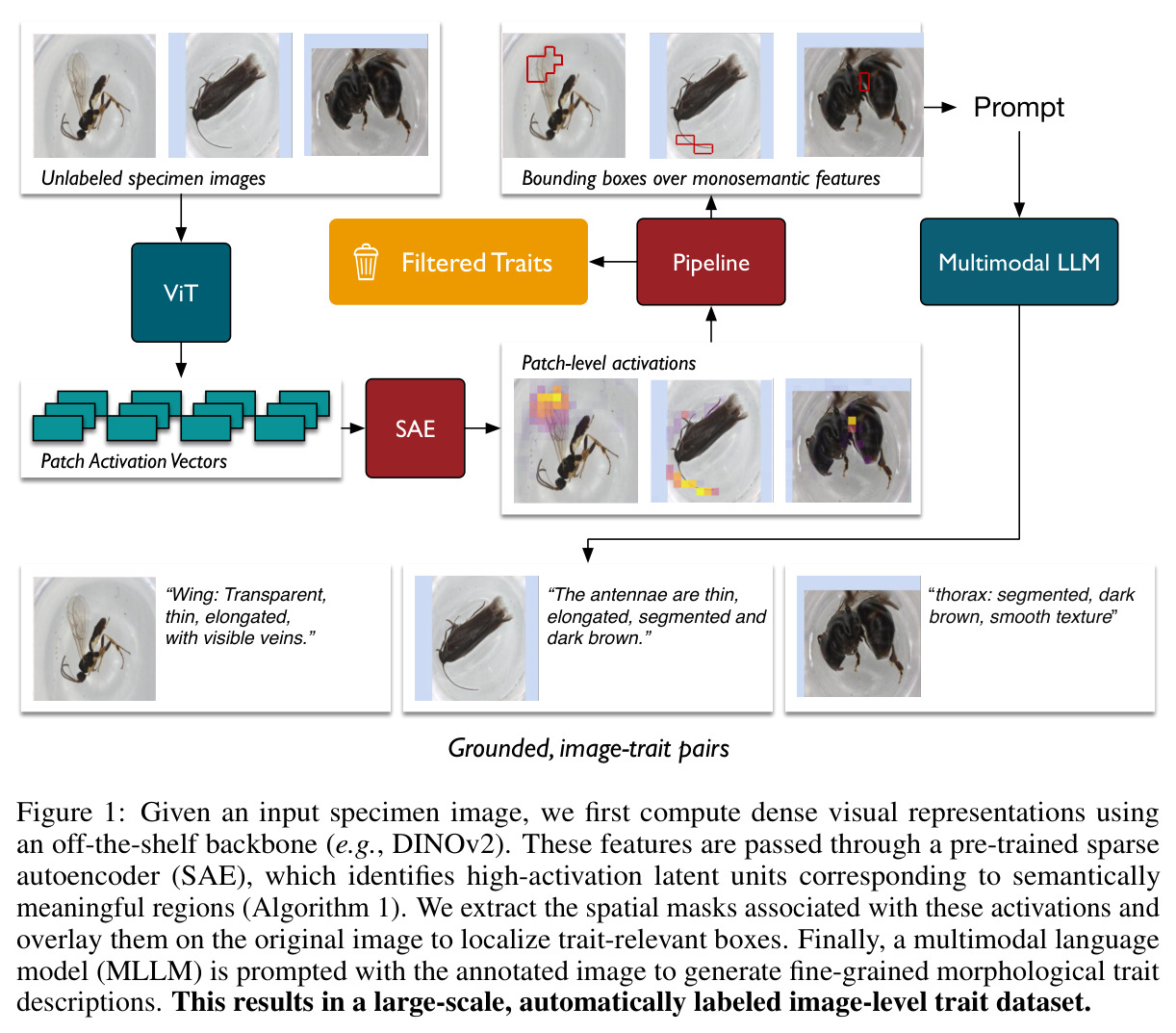
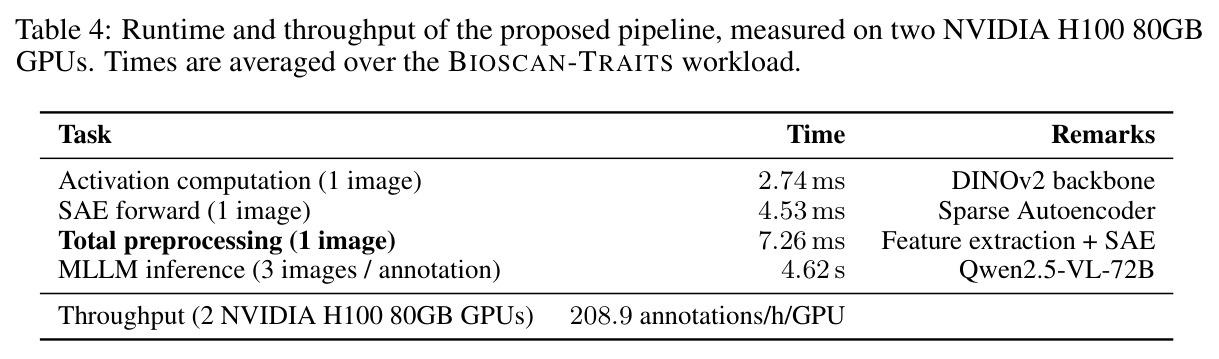
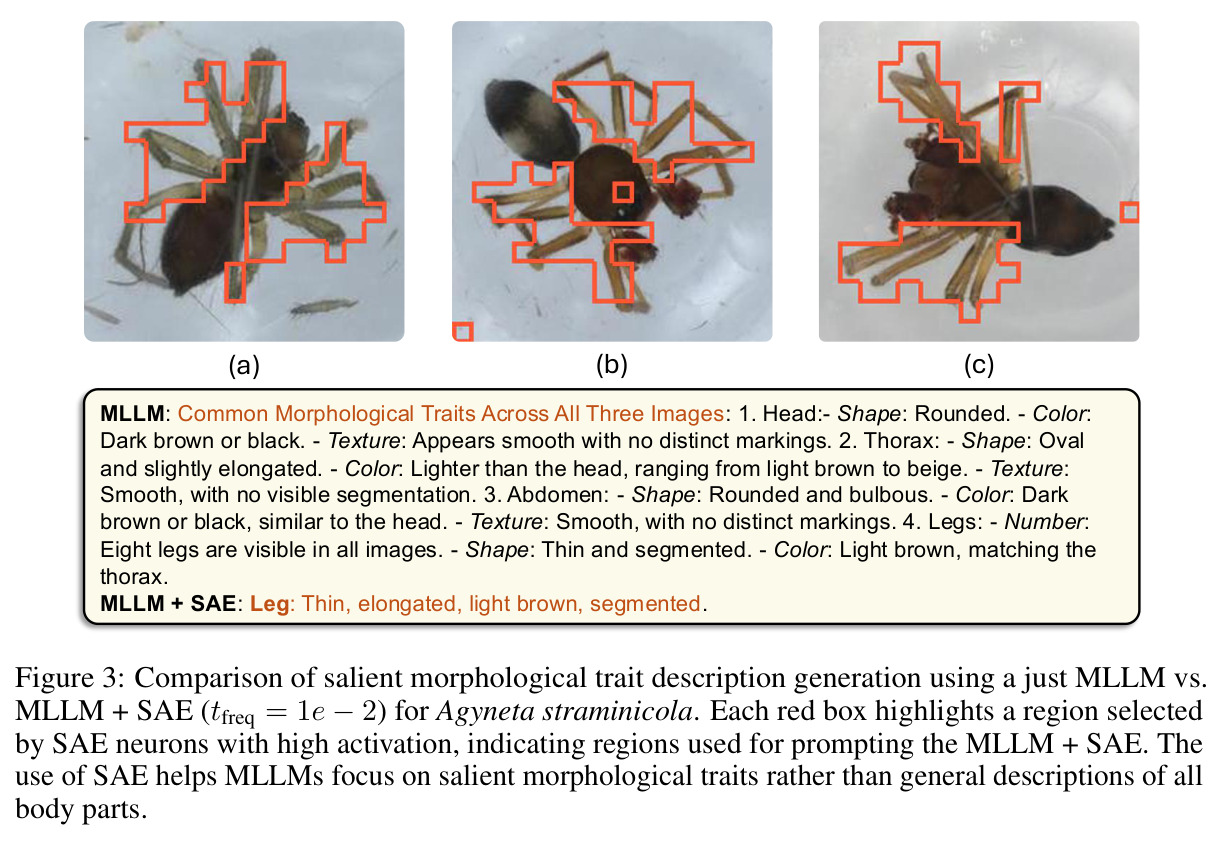
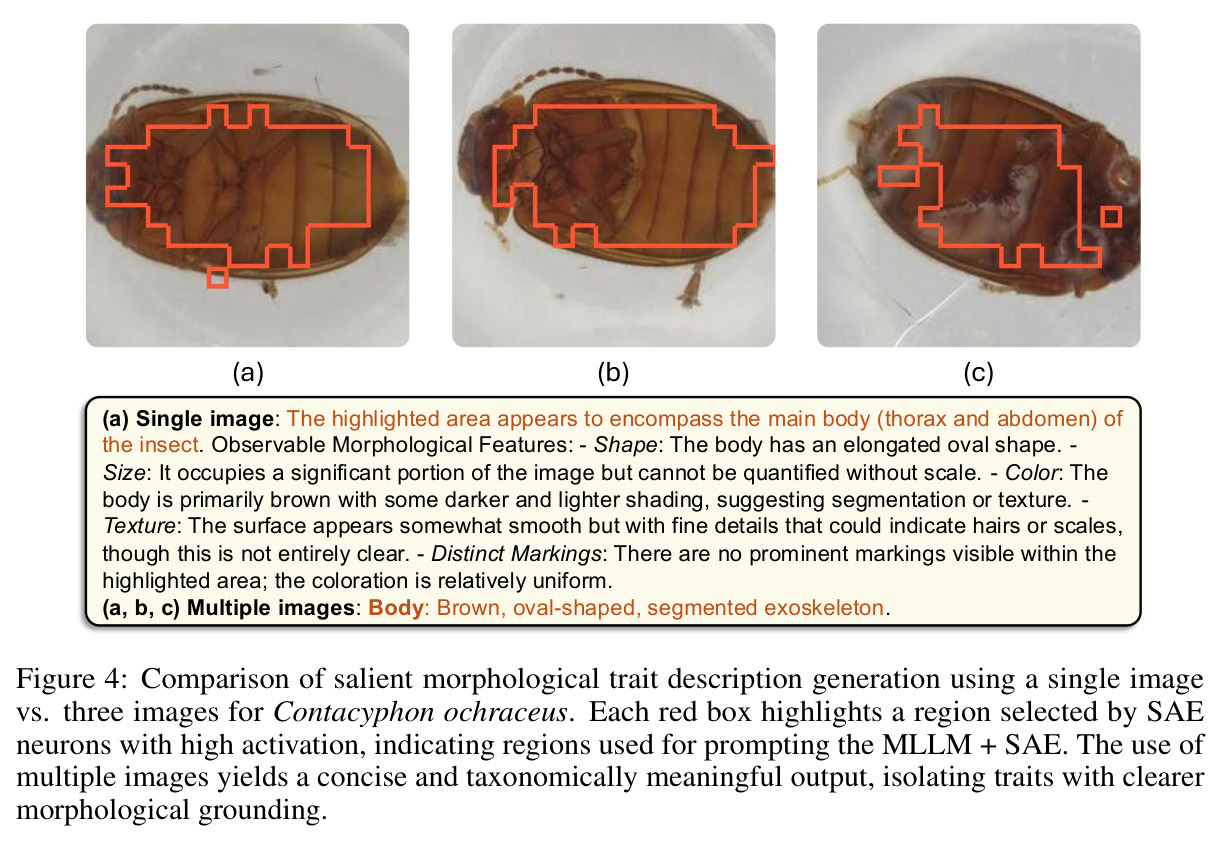
直觉上,可以把 DINOv2 比作一台能拍出'特征照片'的相机,但所有部位都纠缠在一起。SAE 就是一组滤镜,每个滤镜只对某一种语义部件(如'昆虫翅膀')响应,且响应区域在空间上紧致。本文把这种'monosemantic 的稀疏神经元'当作免费的部件提议器:先用 DINOv2 抽 patch 特征 → 训练 ReLU SAE → 统计每个物种和属中各 latent 的激活频率 → 用 species-contrastive 排序挑出'该物种强、其同属近缘物种弱'的诊断性单元 → 把这些单元对应的空间 mask 抠成 bounding box → 用模板 prompt 让 Qwen2.5-VL-72B 针对每个 box 写一段'形状-颜色-纹理'描述 → 多张同种图像取共识,进一步过滤单图噪声。整个流程中,MLLM 不再被要求'描述整张图',而只需'描述这个红框区域',幻觉和背景泄漏大幅下降。
本质创新是把 SAE 当作无监督、可解释、空间定位的部位检测器,并通过物种对比排序(focal species 强 vs. 同属近缘弱)让其天然聚焦于分类学上有诊断价值的部位。与 Grad-CAM(弥散热图、跨物种共享)相比,SAE 神经元是 monosemantic 的(一个神经元对应一种部位);与有监督部件检测器相比,SAE 不需要任何部件级标注;与纯 MLLM 方法相比,SAE 把'描述整图'降级为'描述局部框',让 MLLM 的任务难度从开放域骤降到封闭域。这是把 mechanistic interpretability 工具迁移到生物视觉的最早系统化尝试之一。
方法步骤详情
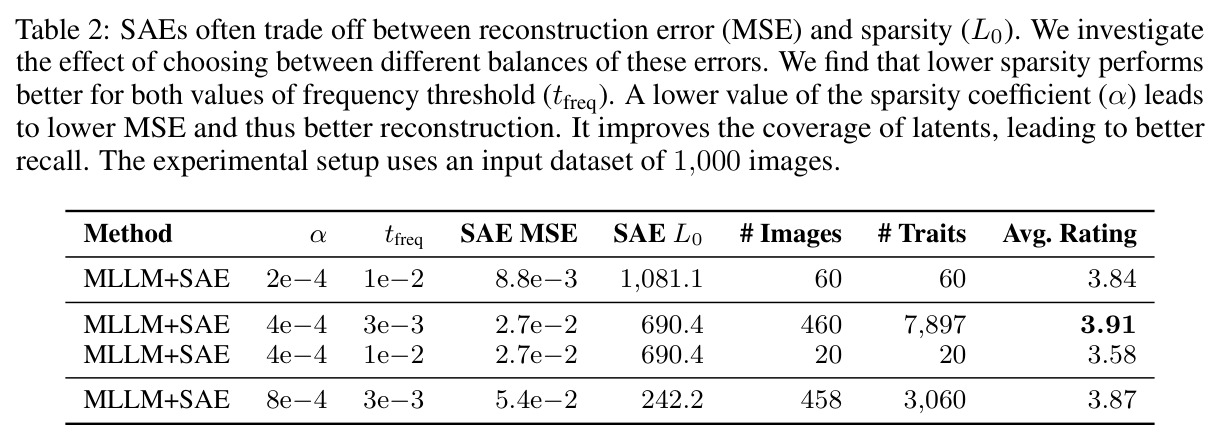
1. **特征抽取**:用 DINOv2-base (ViT-B/14) 倒数第二层(layer 10)对 BIOSCAN-5M 每张图像抽 patch-level 密集特征 z ∈ ℝᵈ。 2. **SAE 训练**:训练一个隐藏宽度 24,576(32× 扩展)的 ReLU SAE,编码器 $u = W_e(z - b_d) + b_e$,稀疏码 $g(z) = \text{ReLU}(u)$,重建 $\tilde{z} = W_d g(z) + b_d$,损失 $\mathcal{J}(\phi) = \|z - \tilde{z}\|_2^2 + \alpha R(g(z))$。稀疏系数 α 在 {2e-4, 4e-4, 8e-4} 之间消融。训练 11 小时,使用 16,384 batch size、学习率 1e-3。 3. **稀疏激活计算 + 阈值筛选**:对每个物种标注样本计算稀疏码,仅保留激活值超过 $t_{activation} = 0.9$ 的 latent。 4. **分类频率聚合**:统计每个 latent 在物种级 $C_{species}[s][z]$ 和属级 $C_{genus}[g][z]$ 的出现频次,计算归一化频率 $f_s(z) = C_{species}[s][z] / \sum_{z'} C_{species}[s][z']$。 5. **Species-Contrastive 筛选**:保留满足 $f_s(z) > t_{freq}$、$f_g(z) > t_{freq}$ **且** $f_s(z) > f_g(z)$ 的 latent——即在该物种中显著高于同属水平。$t_{freq}$ 在 {3e-3, 6e-3, 1e-2} 之间消融。 6. **空间定位**:每个被选中的 latent 都对应一组高激活 patch,把这些 patch 聚成 bounding box。 7. **MLLM 特征描述**:把红框标注的图像输入 Qwen2.5-VL-72B,prompt 要求'识别红框中的身体部位 → 描述形状/大小/颜色/纹理/标记'。多张同种图像取共识,最终每张图像平均生成 4.2 条特征描述。 8. **数据集产出**:在 19K 张物种标注图像上产出 80,808 条特征描述,构成 BIOSCAN-TRAITS。
技术新颖性
技术上,本文最显著的创新是'把 SAE 从 mechanistic interpretability 工具'跨界到'无监督部位检测器'。在 NLP 社区 SAE 主要用于分析 transformer 内部激活(如 Anthropic 的 Claude 3 Sonnet 单义性研究),本文是较早把它系统化地用于生物学图像的工作。方法上,species-contrastive ranking($f_s > f_g$ 的条件)是一个简单但关键的工程创新,它把 SAE 的'通用部位检测'升级为'分类学诊断性部位检测',这在以往 SAE 文献中没有出现过。流水线上,'SAE 定位 → MLLM 描述'的解耦设计让两个模块可以独立替换,实用性大大强于端到端黑盒方案。
实验结果
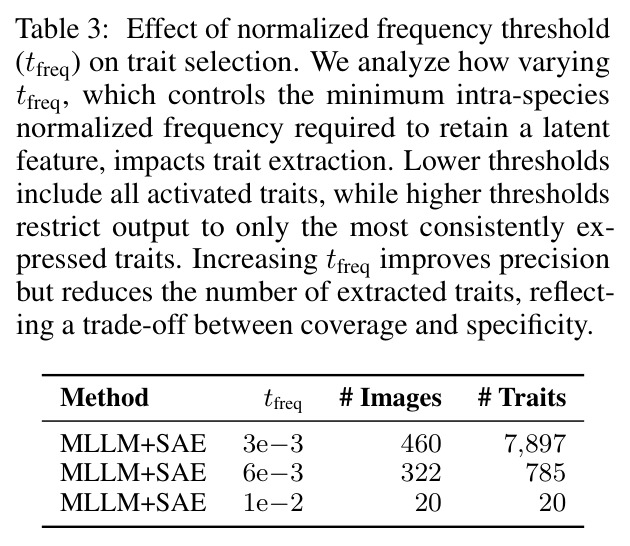
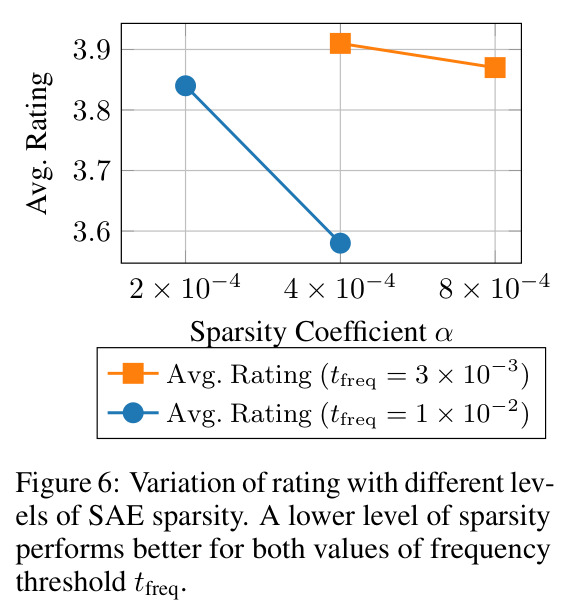
**核心结论 1:SAE 神经元确实 monosemantic 地对应生物部位。** 第 4.4 节展示了 neuron 4852 持续激活在翅膀、neuron 13860 持续激活在触角、neuron 4040 持续激活在胸部、neuron 16584 响应在腿-体连接处、neuron 14153 响应在腹部——这些都是不同目、科的昆虫(鞘翅目、双翅目、膜翅目、缨翅目等),说明 SAE 学到的是真正跨分类群可复用的部位概念。 **核心结论 2:SAE 引导显著提升特征描述质量。** 在 Qwen2.5-VL-72B + 1000 张图像 + $t_{freq}=3\times10^{-3}$ 配置下,MLLM-only(3 图)平均评分 3.15(±0.54),MLLM+SAE(1 图)跃升到 3.84(±0.63),MLLM+SAE(3 图)进一步到 3.91(±0.92)。相对提升约 24%。 **核心结论 3:多图共识机制有效。** MLLM+SAE 从 1 图到 3 图,评分从 3.84 → 3.91,但 token 量从 411 → 1,072(成本上升),特征数从 9,435 → 7,897(更精确过滤)。说明多图约束让模型聚焦'跨实例稳定特征',减少单图特异描述。 **核心结论 4:低稀疏度更好。** α=2e-4(L₀≈1081)评分 3.84;α=4e-4(L₀≈690)评分 3.91;α=8e-4(L₀≈242)评分 3.87($t_{freq}=3\times10^{-3}$)。结论是覆盖更广的潜在部件提议对下游特征提取更友好。 **核心结论 5:$t_{freq}$ 是精度-召回旋钮。** $t_{freq}=3\times10^{-3}$ 保留 7,897 条特征;$t_{freq}=6\times10^{-3}$ 仅 785 条;$t_{freq}=10^{-2}$ 仅 20 条。低阈值保留广覆盖(更多噪声),高阈值只保留跨样本稳定特征。 **核心结论 6:MLLM 规模决定可靠性。** Qwen-2.5-VL-7B 仅 2.90(MLLM+SAE),Qwen-2.5-VL-72B 提升到 3.58,GPT-5-mini 进一步达 4.04(±0.45,方差最小)。一个有趣案例:72B 正确识别红框为'背景',7B 却幻觉出'翅膀'描述。 **核心结论 7:BIOSCAN-TRAITS 显著提升下游分类。** 在 Insects(Ullah et al. 2022)零样本分类基准上,BioCLIP 基线 34.8% → 加物种级微调 39.6% → 加特征级微调 39.9%;BioCLIP 2 基线 55.3% → 加特征级微调 56.23%。这是'机器生成的特征描述'作为弱监督信号首次在该基准上产生可测量的下游收益。



查看结构化数据
| 任务 | 指标 | 本文 | 基线 | 提升 |
|---|---|---|---|---|
| 特征描述人工评分(5 分制,3 标注员均值归一化) | Avg. Rating | MLLM+SAE 3 图: 3.91 (±0.92),单图: 3.84 (±0.63) | MLLM-only 3 图: 3.15 (±0.54),单图: 3.00 (±0.71) | +0.76(24% 相对提升) |
| GPT-5-mini vs Qwen-2.5-VL 系列对比 | Avg. Rating | GPT-5-mini MLLM+SAE: 4.04 (±0.45) | Qwen-2.5-VL-72B MLLM+SAE: 3.58 (±1.05);Qwen-2.5-VL-7B: 2.90 (±1.39) | GPT-5-mini 比开源 72B 高 0.46,比 7B 高 1.14 |
| BioCLIP 零样本物种分类(Insects 基准) | Top-1 Accuracy | BioCLIP+特征微调: 39.9%;BioCLIP 2+特征微调: 56.23% | BioCLIP baseline: 34.8%;BioCLIP 2 baseline: 55.3% | +5.1%(BioCLIP),+0.93%(BioCLIP 2) |
| 特征描述自动生成吞吐量 | Annotations/hour/GPU | 208.9 annotations/h/GPU(双 H100 80GB) | MLLM 推理占 4.62 s/annotation;DINOv2+SAE 仅 7.26 ms/张 | 特征提取几乎免费,主要瓶颈是 MLLM 推理 |
| 特征描述 API 成本 | $/annotation | GPT-5-mini API: $8×10⁻⁴/annotation(100K 张约 $80) | Qwen2.5-VL-72B API (together.ai): $4.1×10⁻³/annotation(100K 张约 $410) | GPT-5-mini 便宜 5.1× 且质量更高 |
| 视觉骨干对比(1000 物种 kNN 分类) | Top-1 Accuracy | DINOv2-base kNN: 41.28% | CLIP ViT-B/16 kNN: 24.57%;SigCLIP ViT-B/16 kNN: 29.68% | DINOv2 比 CLIP 类骨干高 11.6-16.7 个百分点 |
局限与改进
**作者承认的局限**:(1) 假设 DINOv2 特征包含形态相关信号,若骨干偏向通用视觉概念,重要生物学特征可能被遗漏;(2) SAE 单个 latent 可能对应'多个共现特征'(如'细长+薄'),难以解耦组合特征;(3) 7B 级 MLLM 在噪声 patch 上易幻觉;(4) 缺乏大规模真值特征标注,无法在精确率/召回率上做端到端评测;(5) 近期工作(Kantamneni et al. 2025; Wu et al. 2025)指出 SAE 在下游任务上不一定优于简单基线,本文承认但不依赖 SAE 做稀疏 probing,而是把它当作'经过经验验证和过滤的部件提议器'。**我的独立观察**:(a) BIOSCAN-TRAITS 仅覆盖昆虫,对植物/鸟类/真菌的迁移性只是理论承诺,没有跨域实验;(b) SAE 是离线训练一次后冻结的,无法利用新标注增量更新;(c) species-contrastive 排序要求每属至少多个物种才能生效,对单种属(monotypic genus)失效;(d) 数据集 19K 张图仅覆盖 736 物种,平均每物种约 26 张,统计功效有限;(e) 评价全部由作者本人完成(IRB 豁免),存在主观偏差风险。
独立分析的弱点
**弱点 1:缺乏跨分类群验证。** 全部实验都在 BIOSCAN-5M 的昆虫上进行,作者虽然强调方法可迁移到 iNaturalist、TreeOfLife、CUB-200,但没有任何植物/鸟类/真菌的对照实验。一个严谨的补救方案是在 BIOSCAN-TRAITS 同等流程下处理 iNaturalist 的植物子集做零样本迁移测试。 **弱点 2:人工评测规模小且主观。** 每次消融仅随机抽样 30 条特征由 3 位作者打分,存在评分者偏差(虽然做了 per-rater mean 归一化)。改进方向是引入领域生态学家盲评 + 增加每配置样本量到 200+ 并报告 95% 置信区间。 **弱点 3:物种对比排序的边界条件未充分分析。** 当某属只有 1-2 个物种时,$f_s > f_g$ 退化为平凡不等式,方法失效。改进方向是引入'属内最大似然差'或'相对于全数据集的 z-score'作为鲁棒替代指标。 **弱点 4:MLLM 成本是部署瓶颈。** 即使用 GPT-5-mini,100K 图像仍需 $80 和约 480 GPU 小时。改进方向是用 SAE 自身做粗筛,只把真正模糊的 patch 送到 MLLM 仲裁;或蒸馏一个 7B 视觉专家专门做'局部特征描述'任务。 **弱点 5:'monosemantic'是定性观察而非定量度量。** 作者仅展示少数神经元可视化(Figure 5 和 J.11),没有给出'神经元-部位对应'的系统化定量评测。改进方向是用人工标注的部件分割 mask 计算 IoU,或用 probing 任务验证 SAE latent 的语义纯度。
未来方向
**作者提出的方向**:(1) 把流水线扩展到多分类层级(科、属、种),构造跨尺度特征数据库;(2) 应用到非昆虫生物(植物、真菌、鸟类);(3) 用特征级监督训练'领域专用'生物视觉-语言模型。**基于成果可延伸的方向**:(a) 把 BIOSCAN-TRAITS 与 DNA 条形码、地理分布做多模态对齐,做'基因型-表型-环境'联合嵌入;(b) 用 SAE latent 作为可解释性探针,诊断 BioCLIP 在分布外样本上的失败模式;(c) 把 species-contrastive 思想推广到'性别对比''发育阶段对比',生成二态特征(如雌雄异形、性成熟前后形态变化);(d) 与 VLM-as-a-Judge 结合做大规模自动评测,缓解人工评分瓶颈;(e) 把 SAE 替换为更现代的稀疏架构(top-k SAE、Matryoshka SAE)做端到端对比。
复现评估
**开源情况**:代码完全开源在 `github.com/OSU-NLP-Group/sae-trait-annotation`,数据集发布在 `huggingface.co/datasets/osunlp/bioscan-traits`,超参在 Table D.4,prompt 模板在 Appendix C,算法伪代码是 Algorithm 1,可复现性极佳。**算力需求**:SAE 训练约 11 小时(单卡 H100 即可,batch 16,384);数据集生成 193 小时(单进程,2×H100 80GB);下游 BioCLIP 微调未单独说明但量级小。**数据**:BIOSCAN-5M 本身在 NeurIPS 2024 已开放,需注册获取完整版本。**复现难度**:中低。门槛在于 (a) 需要至少 2 张 H100 才能跑通 GPT-5-mini 或 72B 模型的推理;(b) MLLM API 调用有持续成本(100K 图约 $80-410);(c) SAE 训练对显存敏感(24,576 维隐层 + 16,384 batch 至少需 40GB+ 显存)。总体看,**对有 H100 资源的实验室可在 2-3 天内完整复现**;对资源受限的研究者,使用 GPT-5-mini API + 单卡 H100 也是可行方案。
论文图表
补充神经元激活案例:neuron 4040 在胸部、16584 在腿-体连接处、13433 在复眼、14153 在腹部。每个神经元都在多个高阶分类群(瘿蚊科、摇蚊科、蚤蝇科等)上稳定激活。
扩展 Figure 5 的 monosemantic 证据到更多部位和分类群,加强'SAE 学到的是真正可复用的部位概念'这一论断。
三模型对比:7B 把红框背景错误地描述为'翅膀'(幻觉);72B 正确识别为'背景'并拒绝输出;GPT-5-mini 给出更长、更结构化的多部位描述。
直观说明为什么需要 ≥72B 规模的 MLLM,以及为什么 GPT-5-mini 在评分上领先——它有更强的空间定位能力和更少的幻觉。
隐藏宽度 24,576(32× 扩展)、α ∈ {2e-4, 4e-4, 8e-4}、batch size 16,384、学习率 {5e-4, 1e-3}、激活阈值 0.9、ViT layer 10、BioCLIP 微调学习率 3e-4。
工程复现的'唯一可信源',复现时所有超参都需要严格对齐这张表。
736 物种、417 属、19.1K 张唯一图像、80.8K 条特征样本;平均每图 4.2 条特征。
给出数据集的完整规模描述,是评估'数据量是否够下游任务使用'的基础参考。
20K 图像 + tfreq=1e-2 配置下,Qwen-7B MLLM+SAE 2.90、Qwen-72B 3.58、GPT-5-mini 4.04。
给出三个 MLLM 后端在同一条件下的完整对比,是选择部署模型的关键参考。