公平划分颠覆排行榜:CHANRG揭示RNA二级结构预测的泛化局限性 Fair splits flip the leaderboard: CHANRG reveals limited generalization in RNA secondary-structure prediction
CHANRG基准揭示RNA结构预测器在分布外场景下泛化能力严重受限,基础模型优势消失
前置知识
RNA二级结构
RNA分子通过碱基配对形成的局部结构模式,主要由茎区、发卡环、内环、多环和外环等基本单元构成,决定了RNA的三维折叠和生物学功能。RNA二级结构可以用圆点括号表示法或接触矩阵形式表达,是RNA功能预测和药物设计的重要基础。
理解RNA二级结构是本文的核心前提,因为CHANRG基准测试的构建和评估都围绕如何准确预测这些局部结构单元及其连接方式展开。
基础模型
在大规模未标注数据上预训练的神经网络模型,通过学习通用的序列表示来捕获RNA的语言模式。这些模型通常采用Transformer架构,包含数百万到数十亿参数,能够学习序列的上下文依赖关系。在RNA结构预测任务中,基础模型作为编码器提供序列表示,再通过特定的解码器或预测头输出结构预测结果。
本文的核心发现是基础模型在分布外泛化上的局限性,理解基础模型的工作原理对于解读为什么它们在训练集内表现优异但在新场景下失效至关重要。
分布外泛化
模型在与训练数据分布不同的数据上保持性能的能力。在RNA结构预测中,这体现为模型在训练期间未见过的RNA家族、结构架构或基因组背景上准确预测结构的能力。OOD泛化比传统的留出验证更具挑战性,因为它要求模型学习可迁移的结构原理而非记忆训练数据的特定模式。
CHANRG基准测试的核心目标就是评估OOD泛化能力,本文的主要发现就是在严格控制的OOD条件下,不同预测器类别的性能发生了显著反转。
结构化解码器
一类RNA结构预测方法,通过显式的结构约束或优化算法来生成最终结构预测。这些方法通常基于热力学模型(如RNAfold)、统计模型(如CONTRAfold)或混合优化方法,在预测过程中强制执行物理或生物学约束。相比纯数据驱动的方法,结构化解码器在序列到结构映射中引入了领域知识。
本文的关键发现是结构化解码器在OOD场景下比基础模型更鲁棒,这表明显式的结构约束对于泛化到新场景比纯粹的表示学习更为重要。
研究动机
现有的RNA二级结构预测基准测试存在严重局限性,导致对模型泛化能力的评估不准确。首先,广泛使用的bpRNA派生数据集主要基于序列相似性去重,但序列差异较大的RNA仍可能共享高度相似的二级结构拓扑,导致结构冗余样本跨评估边界存在,人为地夸大泛化性能。其次,传统的配对级得分(如F1分数)会掩盖高阶结构错误,包括错误的连接方式和拓扑不匹配。最后,基于基础模型的方法在宽松的基准设置上报告了改进的泛化能力,但这些设置并未充分考虑结构感知、基因组感知和分层OOD评估,因此在更严格的条件下可能无法预测实际的迁移行为。
本文的目标是本文的目标是构建一个严格控制的RNA二级结构预测基准测试,通过结构感知去重、架构感知划分设计和分层评估体系,准确评估不同类别预测器的真实泛化能力。具体而言,CHANRG旨在区分仅对训练数据进行内插的模型和能够泛化到结构新颖RNA家族、clans和基因组稀疏家族的模型,同时提供从碱基对恢复到高阶拓扑重构的多层级评估框架。
与已有工作不同的是,本文的独特切入角度是结合结构感知去重和生物启发的OOD划分设计。传统的基准测试主要关注序列级别的去重和简单的留出验证,而CHANRG同时考虑了序列和结构两个维度的冗余控制,并通过三种生物学上截然不同的OOD regimes(GenA:保留的架构regime;GenC:训练中不存在的clans;GenF:基因组稀疏家族)来全面测试泛化能力。此外,CHANRG引入了分层评估指标体系,从局部碱基对恢复到整体拓扑重构,能够更精细地定位模型失败的模式。
核心方法
CHANRG方法的整体思路是构建一个严格控制的RNA二级结构预测基准测试,通过多阶段数据curator、架构感知划分设计和分层评估体系来准确评估不同预测器类别的泛化能力。技术路线上,首先从Rfam 15.0的10,025,911条序列中通过完整性筛选、高强度序列级去重和基于bpRNA-CosMoS相似性分数的结构感知去重,最终得到170,083个结构非冗余RNA序列。然后基于RNArchitecture定义的分层RNA结构分类方案,结合参考基因组感知规则,构建了留出的同分布Validation和Test划分,以及三个生物学上独特的OOD regimes。最后,在多层级指标体系下评估29个预测器,包括8个结构化解码器、3个直接神经预测器和6个基础模型预测器。
CHANRG的核心创新点是结构感知去重和架构感知划分设计的结合。传统的基准测试仅基于序列相似性去重,但序列差异较大的RNA仍可能共享高度相似的二级结构拓扑,导致结构冗余样本跨评估边界存在。CHANRG通过bpRNA-CosMoS相似性分数进行结构感知去重,即使在高强度序列级过滤后,结构感知修剪仍移除了额外的33倍残差冗余。架构感知划分设计则基于非编码RNA的层级组织结构,结合参考基因组感知规则,将development和evaluation在家族内分离,创建了GenA(保留架构)、GenC(训练中不存在的clans)和GenF(基因组稀疏家族)三种OOD regimes,全面测试不同类型的结构新颖性。
方法步骤详情
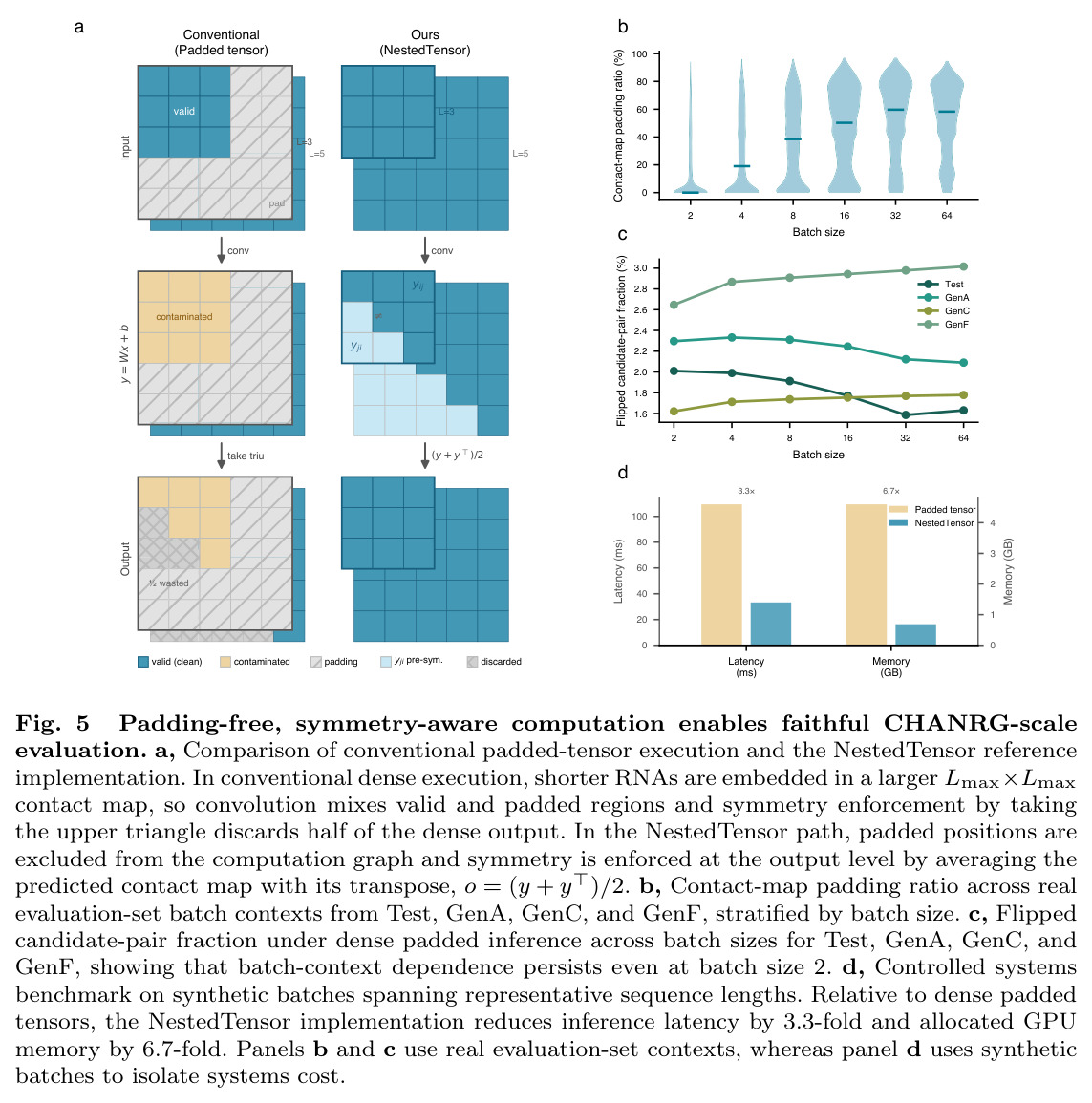
CHANRG的构建过程分为四个主要步骤。第一步是数据收集和完整性筛选,从Rfam 15.0版本收集10,025,911条序列,涉及4,178个源家族,通过完整性筛选后保留10,025,740条序列。第二步是高强度序列级去重,移除序列相似度超过99%的重复序列,得到5,670,054条序列。第三步是基于bpRNA-CosMoS相似性分数的结构感知去重,移除结构相似度超过0.9的冗余序列,最终得到170,083个结构非冗余RNA序列。第四步是架构感知划分设计,基于RNArchitecture定义的分层RNA结构分类方案,结合参考基因组感知规则,将序列分配到Train(123,223条)、Validation(14,070条)、Test(14,070条)和三个OOD regimes(GenA:12,499条,GenC:4,424条,GenF:1,797条)。最后,在多层级指标体系下评估29个预测器,使用padding-free、symmetry-aware参考实现确保评估的batch不变性。
技术新颖性
CHANRG的技术新颖性体现在三个方面。首先,结构感知去重是前所未有的,传统的基准测试主要基于序列级别去重,而CHANRG同时考虑了序列和结构两个维度的冗余控制,结构感知修剪移除了额外的33倍残差冗余。其次,架构感知划分设计是生物启发的,基于非编码RNA的层级组织结构,结合参考基因组感知规则,创建了三种生物学上截然不同的OOD regimes,分别测试保留架构、训练中不存在的clans和基因组稀疏家族的泛化能力。最后,分层评估指标体系是多尺度的,从base-pair F1(局部接触恢复)到stem F1(螺旋级恢复)到topology F1(高阶结构组织)再到topology GED(结构编辑距离),能够更精细地定位模型失败的模式。
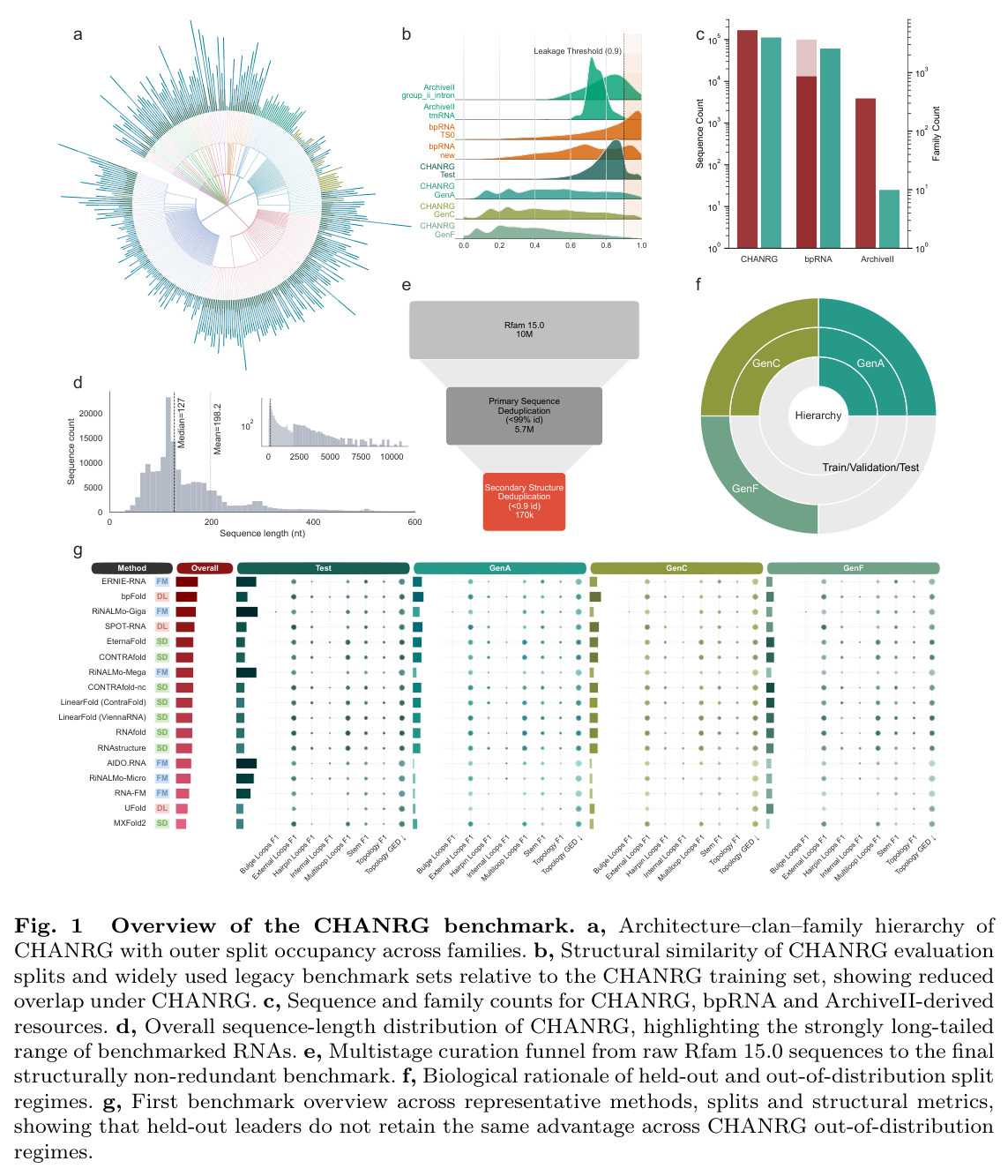
实验结果
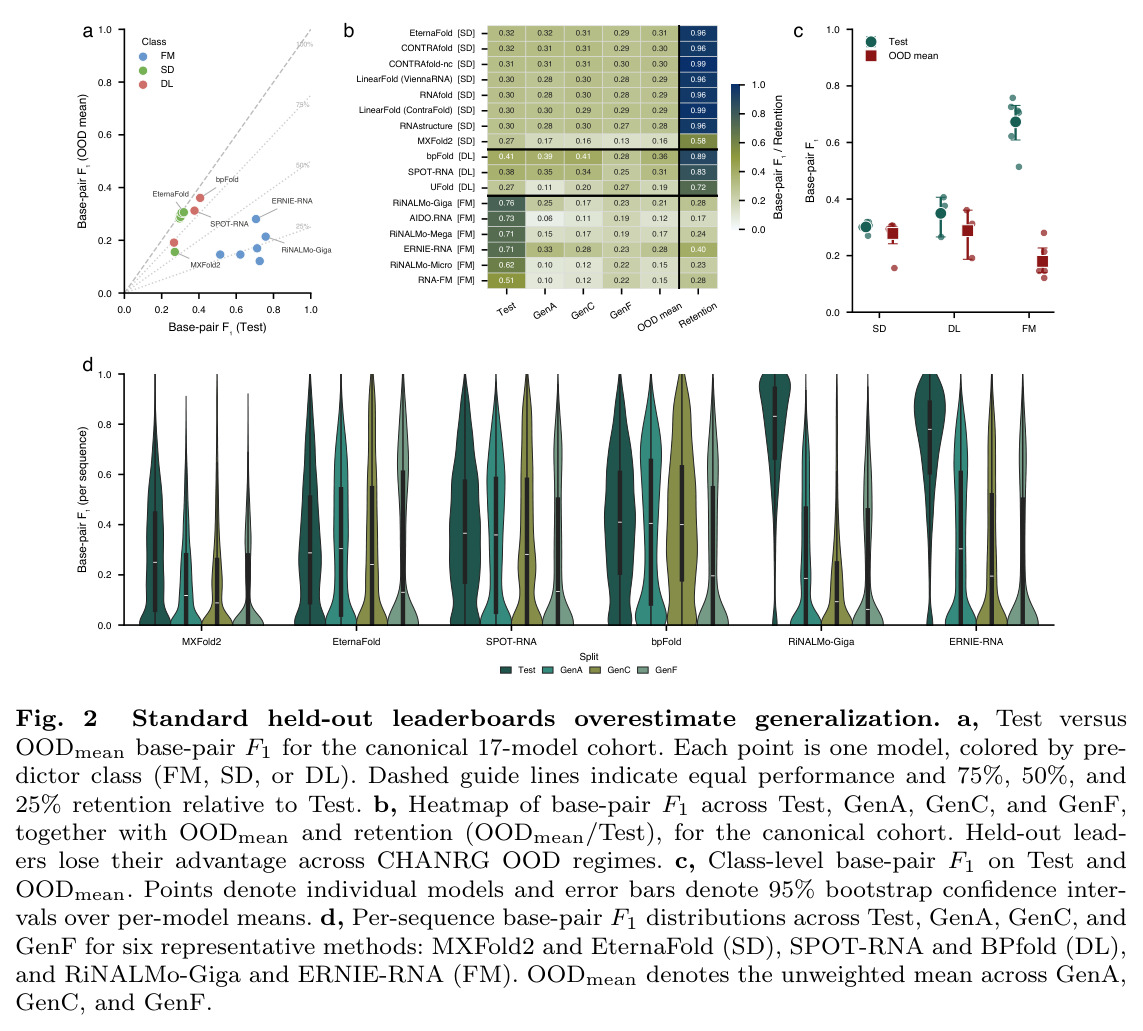
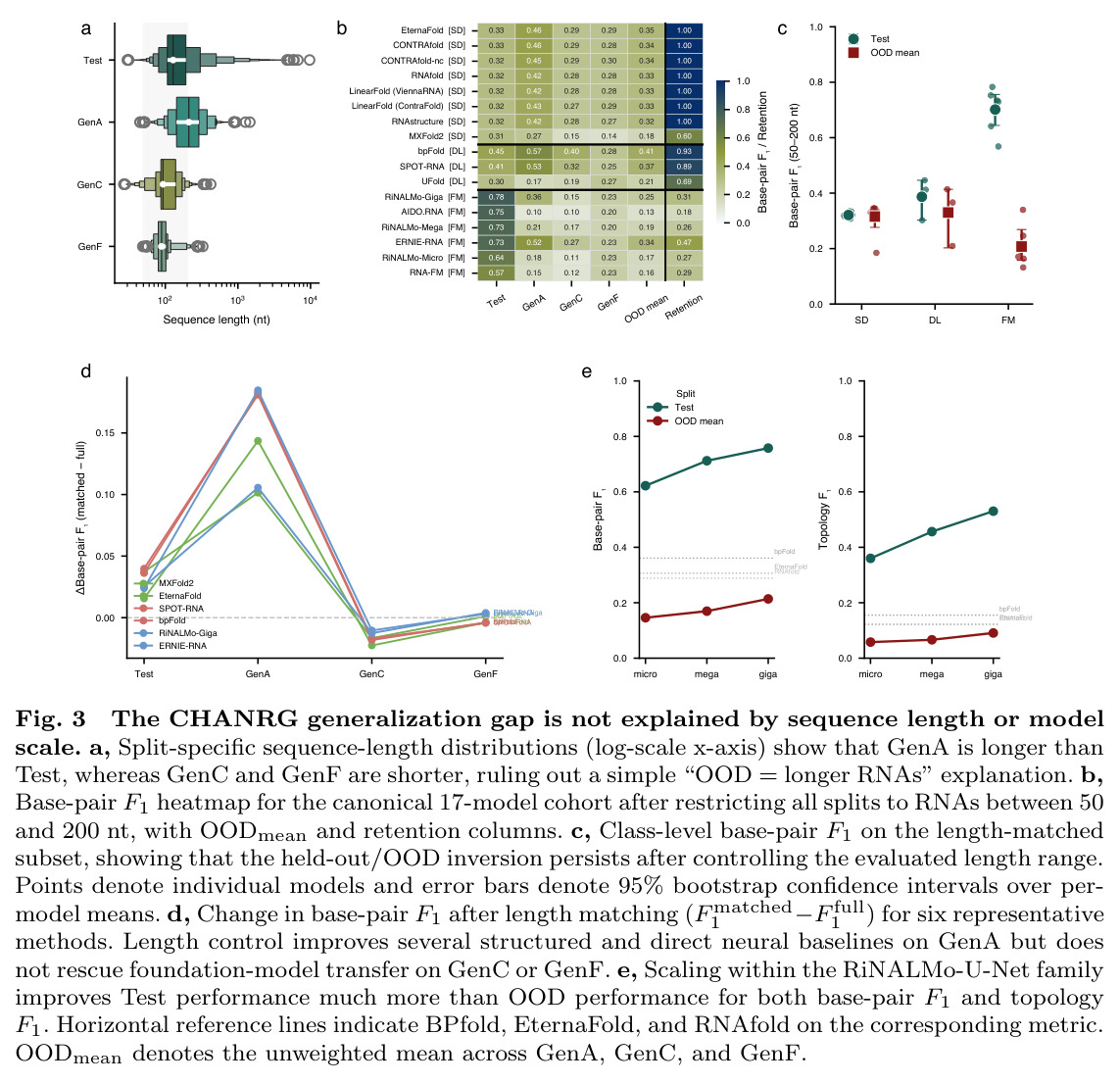
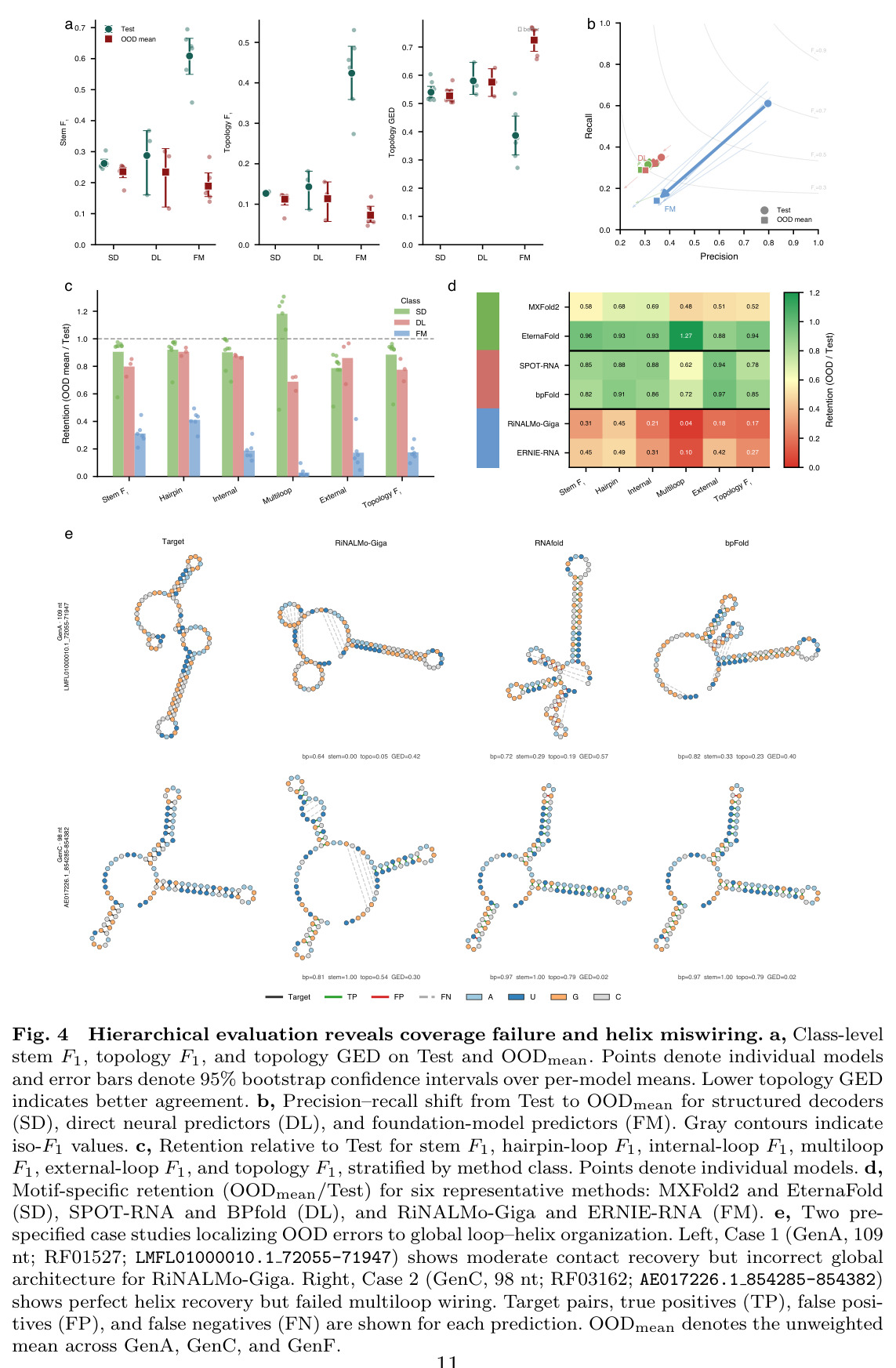
CHANRG基准测试的核心发现是在标准留出leaderboard上表现最佳的预测器并不能最好地泛化到生物学上独特的OOD regimes。在29个预测器的评估中,基础模型预测器在Test集上实现了最高的base-pair F1(0.6731),但在OODmean上仅保留了26.7%的性能(0.1796),对应的绝对损失为0.4935。相比之下,直接神经预测器保留了82.5%的性能(从0.3495下降到0.2883),结构化解码器保留了92.3%的性能(从0.3015下降到0.2784)。在基础模型类别中,RiNALMo-giga-U-Net实现了最高的Test base-pair F1(0.7579),但其在GenA上下降到0.2509,在GenC上下降到0.1651,在GenF上下降到0.2260,总体OODmean为0.2140。在结构化解码器中,EternaFold实现了最强的整体OOD性能,Test base-pair F1为0.3189,OODmean为0.3064。在直接神经预测器中,BPfold实现了最强的整体OOD性能,Test得分为0.4065,OODmean为0.3608。分层评估揭示了两种不同的失败模式:覆盖率失败(遗漏真实相互作用)和连接失败(恢复的螺旋组装成错误的全局架构)。对于RiNALMo-giga-U-Net,pair exact match从Test上的0.0904下降到OODmean上的0.0009,stem F1从0.6947下降到0.2157,topology F1从0.5304下降到0.0910,topology GED从0.2722恶化到0.6570。控制序列长度(50-200 nt)并不能缩小泛化差距,在长度匹配下,基础模型预测器的Test base-pair F1为0.7016,但OODmean仅为0.2074。扩大基础模型规模主要提高留出准确性而非OOD鲁棒性,从micro到giga的scale将Test base-pair F1提高了0.1357,但OODmean仅提高了0.0680。系统基准测试显示,padding-free实现将推理延迟降低了3.3倍,GPU内存分配降低了6.7倍。
查看结构化数据
| 任务 | 指标 | 本文 | 基线 | 提升 |
|---|---|---|---|---|
| RNA二级结构预测(Test集) | Base-pair F1 | RiNALMo-giga-U-Net: 0.7579(基础模型类别平均值:0.6731) | BPfold(直接神经预测器):0.4065,EternaFold(结构化解码器):0.3189 | 最佳基础模型相比最佳基线提升86.4%(0.7579 vs 0.4065) |
| RNA二级结构预测(OODmean) | Base-pair F1 | BPfold(直接神经预测器):0.3608,EternaFold(结构化解码器):0.3064 | RiNALMo-giga-U-Net(基础模型):0.2140(基础模型类别平均值:0.1796) | 最佳基线相比最佳基础模型提升68.6%(0.3608 vs 0.2140) |
| 拓扑结构重构(Test集) | Topology F1 | RiNALMo-giga-U-Net: 0.5304(基础模型类别平均值:0.4240) | BPfold(直接神经预测器):0.1431,EternaFold(结构化解码器):0.1269 | 最佳基础模型相比最佳基线提升270.7%(0.5304 vs 0.1431) |
| 拓扑结构重构(OODmean) | Topology F1 | BPfold(直接神经预测器):0.1136,EternaFold(结构化解码器):0.1125 | RiNALMo-giga-U-Net(基础模型):0.0910(基础模型类别平均值:0.0729) | 最佳基线相比最佳基础模型提升24.8%(0.1136 vs 0.0910) |
| 结构编辑距离(Test集) | Topology GED(越低越好) | RiNALMo-giga-U-Net: 0.2722(基础模型类别平均值:0.3869) | EternaFold(结构化解码器):0.5401,BPfold(直接神经预测器):0.5803 | 最佳基础模型相比最佳基线降低49.6%(0.2722 vs 0.5401) |
| 结构编辑距离(OODmean) | Topology GED(越低越好) | EternaFold(结构化解码器):0.5270,BPfold(直接神经预测器):0.5756 | RiNALMo-giga-U-Net(基础模型):0.6570(基础模型类别平均值:0.7245) | 最佳基线相比最佳基础模型降低19.7%(0.5270 vs 0.6570) |
| 推理效率(相同U-Net架构) | 延迟降低 | Padding-free实现:3.3倍加速 | 传统padded tensor | 延迟从109.4ms降低到33.3ms |
| 内存使用(相同U-Net架构) | 内存降低 | Padding-free实现:6.7倍内存节省 | 传统padded tensor | GPU内存从4.61GB降低到0.69GB |
局限与改进
CHANRG基准测试存在几个局限性。首先,结构感知去重仅与用于定义它的结构表示一样完整,当前的框架对非交叉二级结构拓扑提供了最强的控制,而假结参与的交互在当前比较评估中仍然难以恢复。其次,基准测试评估通过curated注释和结构新颖性的迁移,而不是通过直接下游生物效用,因此高benchmark鲁棒性可能无法直接转化为实验验证的RNA功能改进。第三,CHANRG是结构泛化的严格压力测试,而不是RNA新颖性的详尽分类,其OOD划分是操作性定义的,捕获架构和进化转移的特定形式,但可能不覆盖所有类型的生物学新颖性。第四,训练集和OOD数据集之间的不平衡可能影响结果的解释,特别是GenF只有1,797条序列,而Test有14,070条序列。最后,虽然29个预测器的评估提供了全面的覆盖,但某些新兴方法或最新的基础模型可能未包含在评估中。
独立分析的弱点
CHANRG基准测试的独立分析弱点包括:首先,假结预测的评估仍然有限,当前的bpRNA-CosMoS结构表示主要关注非交叉二级结构拓扑,假结参与的交互难以精确捕捉,这可能导致对某些类型的结构泛化能力评估不足。其次,基准测试主要关注结构预测的准确性,未直接评估下游生物学效用,如RNA功能预测、药物设计靶点识别或实验设计指导,高benchmark鲁棒性可能无法直接转化为实际应用价值。第三,架构感知划分设计虽然生物学上合理,但GenA、GenC和GenF的定义可能无法覆盖所有类型的生物学新颖性,特别是结构新颖性和序列新颖性的交互作用可能未充分探索。第四,虽然padding-free计算解决了batch依赖性问题,但该方法假设接触矩阵的对称性,这对于某些类型的不对称RNA结构预测可能不适用。第五,基准测试主要关注单序列预测,未考虑协进化信息或多序列比对输入,这可能导致对某些类型的预测器不公平。未来的改进方向包括:扩展假结预测的评估框架,引入下游生物学效用的评估任务,探索更多类型的OOD regimes,设计更灵活的计算框架以支持不对称结构预测,以及纳入多序列比对输入的评估。
未来方向
未来的研究方向包括:首先,将假结预测纳入评估框架,当前的bpRNA-CosMoS结构表示主要关注非交叉二级结构拓扑,假结参与的交互难以精确捕捉,开发更全面的结构表示和评估指标将能够更全面地评估结构泛化能力。其次,开发更强可迁移的结构先验,当前的基础模型pipeline未能稳健地恢复高阶结构,这可能需要将结构感知归纳偏差更深度地集成到基础模型架构或训练目标中。第三,设计更丰富的基准测试任务,当前基准测试主要关注结构预测准确性,未来的基准可以包括功能预测、稳定性预测、相互作用预测等更接近实际应用的任务。第四,建立基准测试鲁棒性与实验验证RNA功能之间的更紧密联系,当前基准测试通过curated注释和结构新颖性评估迁移,未来可以设计直接与实验结果相关联的评估任务。第五,探索更多类型的OOD regimes,当前CHANRG定义了三种OOD regimes,但可能还有其他类型的生物学新颖性值得探索,如结构-功能解耦、环境依赖性结构变化等。第六,开发更灵活的计算框架以支持不对称结构预测和更复杂的结构表示,当前padding-free计算假设接触矩阵的对称性,这可能限制某些类型RNA结构的准确预测。
复现评估
CHANRG基准测试的复现评估总体上是高度可复现的。数据方面,基于公开的Rfam 15.0数据集,包含10,025,911条序列,涉及4,178个源家族,经过完整性筛选、高强度序列级去重和结构感知去重后得到170,083个结构非冗余RNA序列,所有数据都是公开可获取的。代码方面,作者提供了完整的评估框架和padding-free、symmetry-aware参考实现,所有29个预测器都通过MultiMolecule框架实例化,确保了标准化的预处理和评分。实现方面,padding-free计算确保了batch不变性,消除了传统密集填充tensor引入的batch依赖性artifact。算力方面,虽然基础模型预测器(特别是RiNALMo-giga-U-Net)需要大量GPU资源,但结构化解码器和直接神经预测器在标准GPU上即可运行。文档方面,论文提供了详细的实验设置、参数配置和评估协议,其他研究者可以按照相同的设置复现结果。然而,需要注意的是,某些基础模型可能需要特定的许可证或访问权限,这可能会限制完全复现。此外,虽然论文提供了详细的实验设置,但某些超参数调优过程可能未完全公开,这可能会影响结果的精确复现。总体而言,CHANRG基准测试在数据、代码、实现和文档方面都提供了充分的复现支持,是一个高度可复现的研究成果。
论文图表